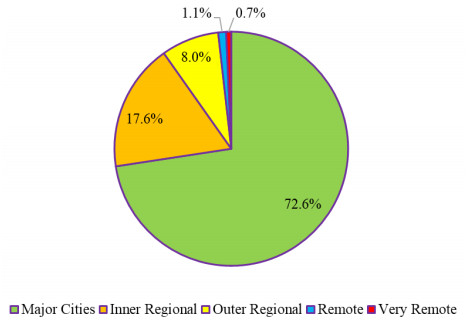
Australian regional universities have been offering many low-enrollment programs to students living in regional, rural, and remote (RRR) areas as part of their obligation to serve regional communities. However, making these programs sustainable has been a significant challenge due to the small population spread across the vast RRR territory in Australia. There is a lack of studies on the key factors contributing to the difficulty of running specific low-enrollment programs, and more importantly, on possible viable solutions to mitigate the negative impact of these factors on program sustainability. Based on the engagement levels of students in teaching and learning, as well as their performance in solving selected questions in basic trigonometry and triangles, I analyzed three independent groups of students enrolled in three offerings of a foundation mathematics course within a low-enrollment secondary mathematics teaching specialty at an Australian regional university before and after the COVID-19 pandemic. Through statistical analysis, I found that: 1) Relaxing entry requirements for the secondary mathematics specialty did not increase student intake post-pandemic; 2) there was no significant difference in performance between active students before and after the pandemic in solving the selected questions under similar teaching and learning conditions; and 3) some students' unsatisfactory performances in solving the selected questions may have been influenced by learning habits inherited from their secondary education, characterized by 'shallow teaching and learning.' This case study also proposes two new ideas, one focused on a single institution and the other on multiple institutions to make this low-enrollment program sustainable at Australian regional universities.
Citation: William Guo. A comparative case study of a foundation mathematics course for student mathematics teachers before and after the COVID-19 pandemic and implications for low-enrollment programs at regional universities[J]. STEM Education, 2025, 5(3): 333-355. doi: 10.3934/steme.2025017
Australian regional universities have been offering many low-enrollment programs to students living in regional, rural, and remote (RRR) areas as part of their obligation to serve regional communities. However, making these programs sustainable has been a significant challenge due to the small population spread across the vast RRR territory in Australia. There is a lack of studies on the key factors contributing to the difficulty of running specific low-enrollment programs, and more importantly, on possible viable solutions to mitigate the negative impact of these factors on program sustainability. Based on the engagement levels of students in teaching and learning, as well as their performance in solving selected questions in basic trigonometry and triangles, I analyzed three independent groups of students enrolled in three offerings of a foundation mathematics course within a low-enrollment secondary mathematics teaching specialty at an Australian regional university before and after the COVID-19 pandemic. Through statistical analysis, I found that: 1) Relaxing entry requirements for the secondary mathematics specialty did not increase student intake post-pandemic; 2) there was no significant difference in performance between active students before and after the pandemic in solving the selected questions under similar teaching and learning conditions; and 3) some students' unsatisfactory performances in solving the selected questions may have been influenced by learning habits inherited from their secondary education, characterized by 'shallow teaching and learning.' This case study also proposes two new ideas, one focused on a single institution and the other on multiple institutions to make this low-enrollment program sustainable at Australian regional universities.

| [1] | Australian Bureau of Statistics, Population clock and pyramid. Commonwealth of Australia, 2024. |
| [2] | Australian Bureau of Statistics, Regional population. Commonwealth of Australia, 2023. Available from: https://www.abs.gov.au/statistics/people/population/regional-population/latest-release. |
| [3] | Australian Institute of Health and Welfare, Rural, regional and remote health: A guide to remoteness classifications. AIHW PHE 53, Canberra, 2004. |
| [4] | Australian Institute of Health and Welfare, Profile of Australia's population, 2024. Retrieved from: https://www.aihw.gov.au/reports/australias-health/profile-of-australias-population. |
| [5] | Regional Universities Network (RUN), RUN Statistics. Australia, 2022. Retrieved from: https://www.run.edu.au/about-us/statistics/ |
| [6] | Nous Group, Economic Impact of the Regional Universities Network. Australia, 2020. |
| [7] | Deloitte Access Economics, Mining and METS: engines of economic growth and prosperity for Australians. Report for the Minerals Council of Australia, 2017. |
| [8] | Wilson, S., Lyons, T. and Quinn, F., Should I stay or should I go? Rural and remote students in first year university stem courses. Australian and International Journal of Rural Education, 2013, 23(2): 77-88. |
| [9] | Fraser, S., Beswick, K. and Crowley, S., Responding to the demands of the STEM education agenda: The experiences of primary and secondary teachers from rural, regional and remote Australia. Journal of Research in STEM Education, 2019, 5(1): 40-59. |
| [10] |
Allen, K.A., Cordoba, B.G., Parks, A. and Arslan, G., Does socioeconomic status moderate the relationship between school belonging and school-related factors in Australia? Child Indicators Research, 2022, 15(5): 1741-1759. https://doi.org/10.1007/s12187-022-09927-3 doi: 10.1007/s12187-022-09927-3

|
| [11] |
Guo, W. and Li, W., A workshop on social media apps for Year-10 students: An exploratory case study on digital technology education in regional Australia. Online Journal of Communication and Media Technologies, 2022, 12(4): e202222. https://doi.org/10.30935/ojcmt/12237 doi: 10.30935/ojcmt/12237
|
| [12] | Ferguson, H., Regional and remote higher education: a quick guide, Commonwealth of Australia, 2022. |
| [13] |
Guo, W., Design and implementation of multi-purpose quizzes to improve mathematics learning for transitional engineering students. STEM Education, 2022, 2(3): 245-261. https://doi.org/10.3934/steme.2022015 doi: 10.3934/steme.2022015
|
| [14] |
Thatcher, A., Zhang, M., Todoroski, H., Chau, A., Wang, J. and Liang, G., Predicting the Impact of COVID-19 on Australian Universities. Journal of Risk and Financial Management, 2020, 13(9): 188. https://doi.org/10.3390/jrfm13090188 doi: 10.3390/jrfm13090188
|
| [15] |
Cook, J., Burke, P.J., Bunn, M. and Cuervo, H., Should I stay or should I go? The impact of the COVID-19 pandemic on regional, rural and remote undergraduate students at an Australian University. Educational Review, 2021, 74(3): 630-644. https://doi.org/10.1080/00131911.2021.1958756 doi: 10.1080/00131911.2021.1958756
|
| [16] | Ngcobo, A.Z., Madonsela, S.P. and Brijlall, D., The teaching and learning of trigonometry. The Independent Journal of Teaching and Learning, 2019, 14(2): 72-91. https://hdl.handle.net/10520/EJC-1d6fb8bc84 |
| [17] |
Fyhn, A.B., What happens when a climber falls? Young climbers mathematise a climbing situation. European Journal of Science and Mathematics Education, 2017, 5(1): 28-42. https://doi.org/10.30935/scimath/9495 doi: 10.30935/scimath/9495
|
| [18] |
Guo, W., Solving word problems involving triangles by transitional engineering students: Learning outcomes and implications. European Journal of Science and Mathematics Education, 2023, 11(2): 249-258. https://doi.org/10.30935/scimath/12582 doi: 10.30935/scimath/12582
|
| [19] |
Dündar, S., Mathematics teacher-candidates' performance in solving problems with different representation styles: The trigonometry example. Eurasia Journal of Mathematics, Science & Technology Education, 2015, 11(6): 1379-1397. https://doi.org/10.12973/eurasia.2015.1396a doi: 10.12973/eurasia.2015.1396a
|
| [20] | Guo, W., Exploratory case study on solving word problems involving triangles by pre-service mathematics teachers in a regional university in Australia. Mathematics, 2022, 10(20): 3786. http://doi.org/10.3390/math10203786 |
| [21] | CQU. (n.d.). Bachelor of Education (Secondary): Mathematics Minor. CQUni Handbook. Retrieved from: https://handbook.cqu.edu.au/he/courses/view/CC13 |
| [22] | Christensen, L.B., Johnson, R.B., Turner, L.A. and Christensen, L.B., Research methods, design, and analysis. Pearson, 2020. |
| [23] | Guo, W.W., Essentials and Examples of Applied Mathematics, 2nd ed. Pearson Australia, 2020. |
| [24] |
Guo W., Solving word problems involving triangles and implications on training pre-service mathematics teachers. STEM Education, 2024, 4(3): 263-281. https://doi.org/10.3934/steme.2024016 doi: 10.3934/steme.2024016
|
| [25] | Preacher, K.J., Calculation for the chi-square test: An interactive calculation tool for chi-square tests of goodness of fit and independence[Computer software], 2001. Available from: http://quantpsy.org. |
| [26] | Sparrow, L. and Swan, P., Calculator use in Western Australian primary schools, Mathematics, Science & Technology Education Centre, Edith Cowan University, Australia, 1997. |
| [27] | Kissane, B., McConney, A. and Ho, K.F., Review of the use of technology in Mathematics education and the related use of CAS calculators in external examinations and in post school tertiary education settings, School Curriculum and Standards Authority, Western Australia, 2015. |
| [28] |
Eleje, L.I., Onuigbo, C.F., Metu, I.C., Ejinkonye, F.O. and Enemouh, U.S., Influence of Secondary School Students' Use of Calculators on their Numerical Skills. Futurity of Social Sciences, 2024, 2(3): 18-40. https://doi.org/10.57125/FS.2024.09.20.02 doi: 10.57125/FS.2024.09.20.02
|




Figures(5) / Tables(15)
William Guo. A comparative case study of a foundation mathematics course for student mathematics teachers before and after the COVID-19 pandemic and implications for low-enrollment programs at regional universities[J]. STEM Education, 2025, 5(3): 333-355. doi: 10.3934/steme.2025017







 DownLoad:
DownLoad: