Bitcoin has become quite known after the 2008 economic crisis and the COVID-19 health crisis. For some, these cryptocurrencies constitute rebellion against the existing system as governments encourage uncontrolled expansions in the money supply; for some others, it is a quick source of income. Undeniably, the volume of the crypto money market has grown considerably in recent years, regardless of the reasoning of the people who invest and trade in this field. At this point, one of the most important questions to be investigated is "what variables have caused the tremendous growth in the crypto money quantities in recent years?" This study tests the assumption that changes in cryptocurrencies are affected by changes in national currencies. Thus, the Bitcoin price is the dependent variable, and M1 monetary supply changes in the USA, European Union and Japanese economies are considered independent variables. The variables in this study were tested using the time-varying Granger causality method. The results obtained from this study confirm the philosophy of Bitcoin's emergence and the possibility that it can be a hedge against the inflationary effects of money, especially after the COVID-19 pandemic.
Citation: Nilcan Mert, Mustafa Caner Timur. Bitcoin and money supply relationship: An analysis of selected country economies[J]. Quantitative Finance and Economics, 2023, 7(2): 229-248. doi: 10.3934/QFE.2023012
Bitcoin has become quite known after the 2008 economic crisis and the COVID-19 health crisis. For some, these cryptocurrencies constitute rebellion against the existing system as governments encourage uncontrolled expansions in the money supply; for some others, it is a quick source of income. Undeniably, the volume of the crypto money market has grown considerably in recent years, regardless of the reasoning of the people who invest and trade in this field. At this point, one of the most important questions to be investigated is "what variables have caused the tremendous growth in the crypto money quantities in recent years?" This study tests the assumption that changes in cryptocurrencies are affected by changes in national currencies. Thus, the Bitcoin price is the dependent variable, and M1 monetary supply changes in the USA, European Union and Japanese economies are considered independent variables. The variables in this study were tested using the time-varying Granger causality method. The results obtained from this study confirm the philosophy of Bitcoin's emergence and the possibility that it can be a hedge against the inflationary effects of money, especially after the COVID-19 pandemic.

| [1] | Acemoglu D, Johnson S, Querubin P (2008) When does policy reform work? The case of central bank independence. Working Paper 14033. National Bureau of Economic Research. https://doi.org/10.3386/w14033 |
| [2] |
Alesina A, Summers LH (1993) Central bank independence and macroeconomic performance: some comparative evidence. J Money Credit Bank 25: 151–162. https://doi.org/10.2307/2077833 doi: 10.2307/2077833

|
| [3] | Arnone M, Laurens B, Segalotto JF, et al. (2007) Central bank autonomy: lessons from global trends. https://doi.org/10.2139/ssrn.986816 |
| [4] |
Balcilar M, Ozdemir ZA, Arslanturk Y (2010) Economic growth and energy consumption causal nexus viewed through a bootstrap rolling window. Energy Econ 32: 1398–1410. https://doi.org/10.1016/j.eneco.2010.05.015 doi: 10.1016/j.eneco.2010.05.015
|
| [5] | Bank of Japan (BOJ) M1 and Components. Available from: https://www.stat-search.boj.or.jp/ssi/cgi-bin/famecgi2?cgi = $nme_a000_en & lstSelection = MD02. |
| [6] |
Benigno P, Schilling LM, Uhlig H (2022) Cryptocurrencies, currency competition, and the impossible trinity. J Int Econ 136: 103601. https://doi.org/10.1016/j.jinteco.2022.103601 doi: 10.1016/j.jinteco.2022.103601
|
| [7] |
Blau BM, Griffith TG, Whitby RJ (2021) Inflation and Bitcoin: A descriptive time-series analysis. Econ Lett 203: 109848. https://doi.org/10.1016/j.econlet.2021.109848 doi: 10.1016/j.econlet.2021.109848
|
| [8] |
Bouri E, Molnár P, Azzi G, et al. (2017a) On the hedge and safe haven properties of Bitcoin: Is it really more than a diversifier? Financ Res Lett 20: 192–198. https://doi.org/10.1016/j.frl.2016.09.025 doi: 10.1016/j.frl.2016.09.025
|
| [9] |
Bouri E, Gupta R, Tiwari AK, et al. (2017b) Does Bitcoin hedge global uncertainty? Evidence from wavelet-based quantile-in-quantile regressions. Financ Res Lett 23: 87–95. https://doi.org/10.1016/j.frl.2017.02.009 doi: 10.1016/j.frl.2017.02.009
|
| [10] | Chohan UW (2021) Cryptocurrencies and Hyperinflation. Critical Blockchain Research Initiative (CBRI) Working Papers. https://dx.doi.org/10.2139/ssrn.3320702 |
| [11] |
Conlon T, Corbet S, Mcgee RJ (2021) Inflation and cryptocurrencies revisited: A time-scale analysis. Econ Lett 206: 109996. https://doi.org/10.1016/j.econlet.2021.109996 doi: 10.1016/j.econlet.2021.109996
|
| [12] | Cukierman A, Kalaitzidakis P, Summers LH, et al. (1993) Central bank independence, growth, investment, and real rates, In: Carnegie-Rochester Conference Series on Public Policy, North-Holland, 39: 95–140. https://doi.org/10.1016/0167-2231(93)90005-H |
| [13] | De Haan J, Eijffinger SC (2016) The Politics of Central Bank Independence. De Nederlandsche Bank Working Paper No. 539. Available at SSRN. http://doi.org/10.2139/ssrn.2888836 |
| [14] |
Dickey DA, Fuller WA (1981) Likelihood ratio statistics for autoregressive time series with a unit root. Econometrica: J Econometric Society, 1057–1072. https://doi.org/10.2307/1912517 doi: 10.2307/1912517
|
| [15] |
Dolado JJ, Lütkepohl H (1996) Making Wald Tests Work for Cointegrated VAR Systems. Econometric Rev 15: 369–386. https://doi.org/10.1080/07474939608800362 doi: 10.1080/07474939608800362
|
| [16] |
Dyhrberg AH (2016) Hedging capabilities of bitcoin. Is it the virtual gold? Financ Res Lett 16: 139–144. https://doi.org/10.1016/j.frl.2015.10.025 doi: 10.1016/j.frl.2015.10.025
|
| [17] | Eğilmez M (2014) Japonya Slumpflasyona Girdi. Kendime Yazılar. Available from: https://www.mahfiegilmez.com/2014/11/japonya-slumpflasyona-girdi.html?m = 1. |
| [18] | European Central Bank (ECB) M1 and Components Available from: https://www.ecb.europa.eu/stats/money_credit_banking/monetary_aggregates/html/index.en.html. |
| [19] |
Fang L, Bouri E, Gupta R, et al. (2019) Does global economic uncertainty matter for the volatility and hedging effectiveness of Bitcoin? Int Rev Financ Anal 61: 29–36. https://doi.org/10.1016/j.irfa.2018.12.010 doi: 10.1016/j.irfa.2018.12.010
|
| [20] | Federal Reserve (FED) M1 and Components. Available from: https://fred.stlouisfed.org/series/M1SL. |
| [21] | Gökalp N (2003) Ekonomide güven faktörü. Yönetim ve Ekonomi Dergisi 10: 163–174. |
| [22] | Greco TH (2009) The End of Money and the Future of Civilization. Vermont: Chelsea Green. ISBN: 978-1-60358-078-6. |
| [23] | Haşlak Ş (2018) Analysis of Bitcoin market volatility. Published master's thesis. Çankaya University, Institute of Social Sciences, Ankara. |
| [24] | Insight Investment (2020) Investment Reports: Covid-19 is Likely to End the Inflation Lockdown. |
| [25] |
Kapetanios G, Shin Y, Snell A (2003) Testing for a unit root in the nonlinear STAR framework. J Econometrics 112: 359–379. https://doi.org/10.1016/S0304-4076(02)00202-6 doi: 10.1016/S0304-4076(02)00202-6
|
| [26] | Kendirkiran G (2021) The relationship between economic growth and foreign trade in Turkey: Time-varying causality approaches. Master's thesis, Ankara Hacı Bayram Veli University, The Institute of Graduate Studies. |
| [27] |
Kostakis V, Giotitsas C (2014) The (A) political economy of bitcoin. Triple C: Communication, Capitalism Critique 12: 431–440. https://doi.org/10.31269/triplec.v12i2.606 doi: 10.31269/triplec.v12i2.606
|
| [28] |
Leybourne SJ (1995) Testing for unit roots using forward and reverse Dickey-Fuller regressions. Oxford B Econ Stat 57: 559–571. https://doi.org/10.1111/j.1468-0084.1995.tb00040.x doi: 10.1111/j.1468-0084.1995.tb00040.x
|
| [29] | Litterman RB (1980) Bayesian procedure for forecasting with vector autoregressions. Massachusetts Institute of Technology. https://doi.org/10.21034/wp.115 |
| [30] |
Litterman RB (1986) Forecasting with Bayesian vector autoregressions—five years of experience. J Bus Econ Stat 4: 25–38. https://doi.org/10.1080/07350015.1986.10509491 doi: 10.1080/07350015.1986.10509491
|
| [31] |
Ma C, Tian Y, Hsiao S, et al. (2022) Monetary policy shocks and Bitcoin prices. Res Int Bus Financ 62: 101711. https://doi.org/10.1016/j.ribaf.2022.101711 doi: 10.1016/j.ribaf.2022.101711
|
| [32] | Nakamoto S (2008) Bitcoin: A peer-to-peer electronic cash system. Available from: https://bitcoin.org/bitcoin. |
| [33] |
Narayan PK, Narayan S, Rahman RE, et al. (2019) Bitcoin price growth and Indonesia's monetary system. Emerg Mark Rev 38: 364–376. https://doi.org/10.1016/j.ememar.2018.11.005 doi: 10.1016/j.ememar.2018.11.005
|
| [34] |
Otero J, Smith J (2012) Response surface models for the Leybourne unit root tests and lag order dependence. Comput Stat 27: 473–486. https://doi.org/10.1007/s00180-011-0268-y doi: 10.1007/s00180-011-0268-y
|
| [35] |
Otero J, Smith J (2017) Response surface models for OLS and GLS detrending-based unit-root tests in nonlinear ESTAR models. Stata J 17: 704–722. https://doi.org/10.1177/1536867X1701700310 doi: 10.1177/1536867X1701700310
|
| [36] |
Perez C (2009) The double bubble at the turn of the century: technological roots and structural implications. Camb J Econ 33: 779–805. https://dx.doi.org/10.1093/cje/bep028 doi: 10.1093/cje/bep028
|
| [37] |
Phillips PC, Shi S, Yu J (2015a) Testing for multiple bubbles: historical episodes of exuberance and collapse in the S & P 500. Int Econ Rev 56: 1043–1078. https://doi.org/10.1111/iere.12132 doi: 10.1111/iere.12132
|
| [38] |
Phillips PC, Shi S, Yu J (2015b) Testing for multiple bubbles: limit theory of real-time detectors Int Econ Rev 56: 1079–1134. https://doi.org/10.1111/iere.12131 doi: 10.1111/iere.12131
|
| [39] | Pirgaip B, Dinçergök B, Haşlak Ş (2019) Bitcoin Market Price Analysis and an Empirical Comparison with Main Currencies, Commodities, Securities and Altcoins, In: Hacioglu, U. (eds) Blockchain Economics and Financial Market Innovation, Contributions to Economics. https://doi.org/10.1007/978-3-030-25275-5_8 |
| [40] | Savaş VF (2016) Politik İktisat (8. baskı). Beta Yayınları, İstanbul. |
| [41] |
Schilling L, Uhlig H (2019a) Some simple bitcoin economics. J Monetary Econ 106: 16–26. https://doi.org/10.1016/j.jmoneco.2019.07.002 doi: 10.1016/j.jmoneco.2019.07.002
|
| [42] | Schilling LM, Uhlig H (2019b) Currency substitution under transaction costs, In: AEA Papers and Proceedings, 109: 83–87. https://doi.org/10.1257/pandp.20191017 |
| [43] |
Shi S, Hurn S, Phillips PC (2020) Causal change detection in possibly integrated systems: Revisiting the money–income relationship. J Financ Econometrics 18: 158–180. https://doi.org/10.1093/jjfinec/nbz004 doi: 10.1093/jjfinec/nbz004
|
| [44] |
Shi S, Phillips PC, Hurn S (2018) Change detection and the causal impact of the yield curve. J Time Series Anal 39: 966–987. https://doi.org/10.1111/jtsa.12427 doi: 10.1111/jtsa.12427
|
| [45] | StataCorp (2021) Stata: Release 17. Statistical Software. College Station, TX: StataCorp LLC. |
| [46] |
Su CW, Qin M, Tao R, et al. (2020) Can Bitcoin hedge the risks of geopolitical events? Technol Forecast Social Change 159: 120182. https://doi.org/10.1016/j.techfore.2020.120182 doi: 10.1016/j.techfore.2020.120182
|
| [47] |
Swanson NR (1998) Money and output viewed through a rolling window. J Monetary Econ 41: 455–474. https://doi.org/10.1016/S0304-3932(98)00005-1 doi: 10.1016/S0304-3932(98)00005-1
|
| [48] |
Thoma MA (1994) Subsample Instability and Asymmetries in Money-Income Causality. J Econometrics 64: 279–306. https://doi.org/10.1016/0304-4076(94)90066-3 doi: 10.1016/0304-4076(94)90066-3
|
| [49] |
Urquhart A, Zhang H (2019) Is Bitcoin a hedge or safe haven for currencies? An intraday analysis. Int Rev Financ Anal 63: 49–57. https://doi.org/10.1016/j.irfa.2019.02.009 doi: 10.1016/j.irfa.2019.02.009
|
| [50] |
Wang L, Sarker PK, Bouri E (2022) Short-and Long-Term Interactions between Bitcoin and Economic Variables: Evidence from the US. Comput Econ, 1–26. https://doi.org/10.1007/s10614-022-10247-5 doi: 10.1007/s10614-022-10247-5
|
| [51] |
Warsito OLD (2020) Analisis Volatilitas Cryptocurrency, Emas, Dollar, Dan Indeks Harga Saham (Ihsg). Int J Social Sci Bus 4: 40–46. https://doi.org/10.23887/ijssb.v4i1.23887 doi: 10.23887/ijssb.v4i1.23887
|
| [52] | Weber B (2014) Can Bitcoin compete with money? J Peer Prod 4. |
| [53] |
Wu S, Tong M, Yang Z, et al. (2019) Does gold or Bitcoin hedge economic policy uncertainty? Financ Res Lett 31: 171–178. https://doi.org/10.1016/j.frl.2019.04.001 doi: 10.1016/j.frl.2019.04.001
|
| [54] | Yahoo Finance. Historical Bitcoin Prices. Available From: https://finance.yahoo.com/quote/BTC-USD?p = BTC-USD & .tsrc = fin-srch. |
| [55] |
Toda HY, Yamamoto T (1995) Statistical inference in vector autoregressions with possibly integrated processes. J Econometrics 66: 225–250. https://doi.org/10.1016/0304-4076(94)01616-8 doi: 10.1016/0304-4076(94)01616-8
|
| [56] |
Yamada H, Toda HY (1998) Inference in possibly integrated vector autoregressive models: Some finite sample evidence. J Econometrics 86: 55–95. https://doi.org/10.1016/S0304-4076(97)00109-7 doi: 10.1016/S0304-4076(97)00109-7
|
| [57] |
Zhu Y, Dickinson D, Li J (2017) Analysis on the influence factors of Bitcoin's price based on VEC model. Financ Innov 3: 1–13. https://doi.org/10.1186/s40854-017-0054-0 doi: 10.1186/s40854-016-0051-8
|
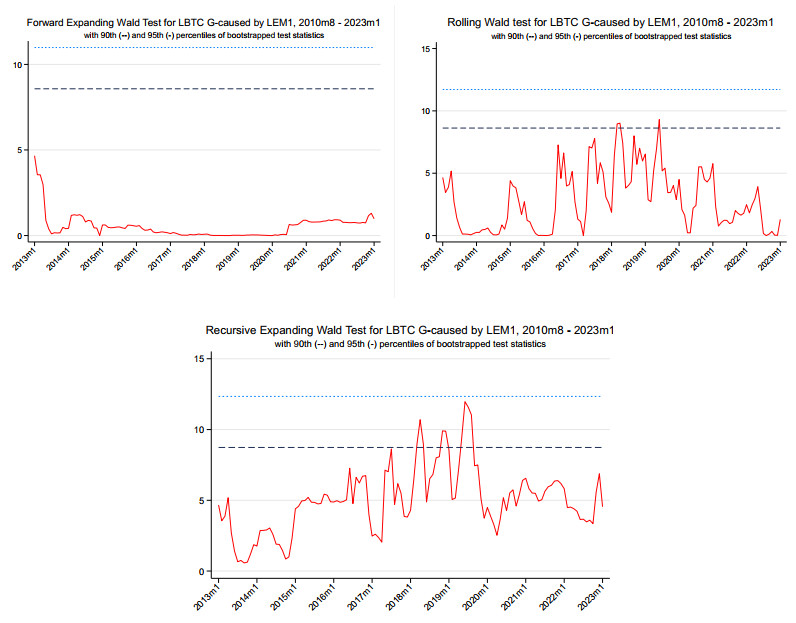
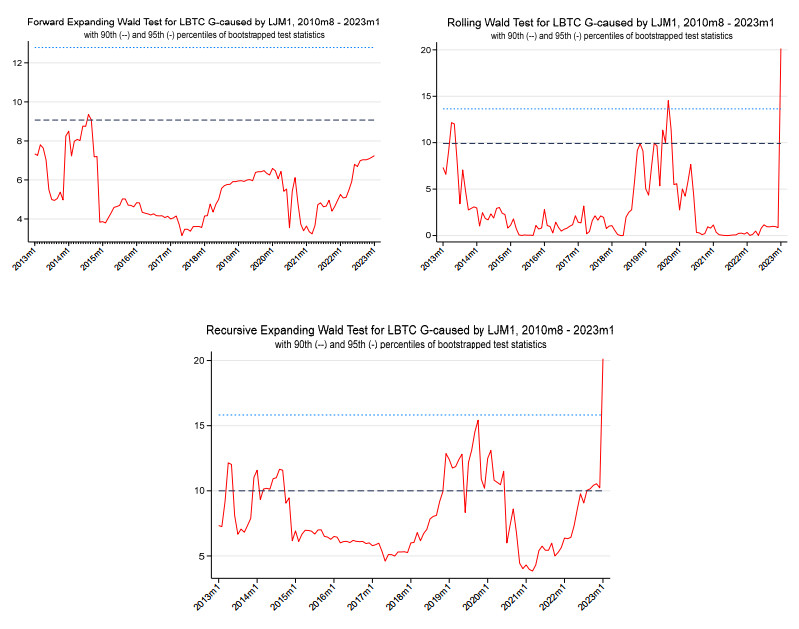
Figures(6) / Tables(7)

Nilcan Mert, Mustafa Caner Timur. Bitcoin and money supply relationship: An analysis of selected country economies[J]. Quantitative Finance and Economics, 2023, 7(2): 229-248. doi: 10.3934/QFE.2023012







 DownLoad:
DownLoad: