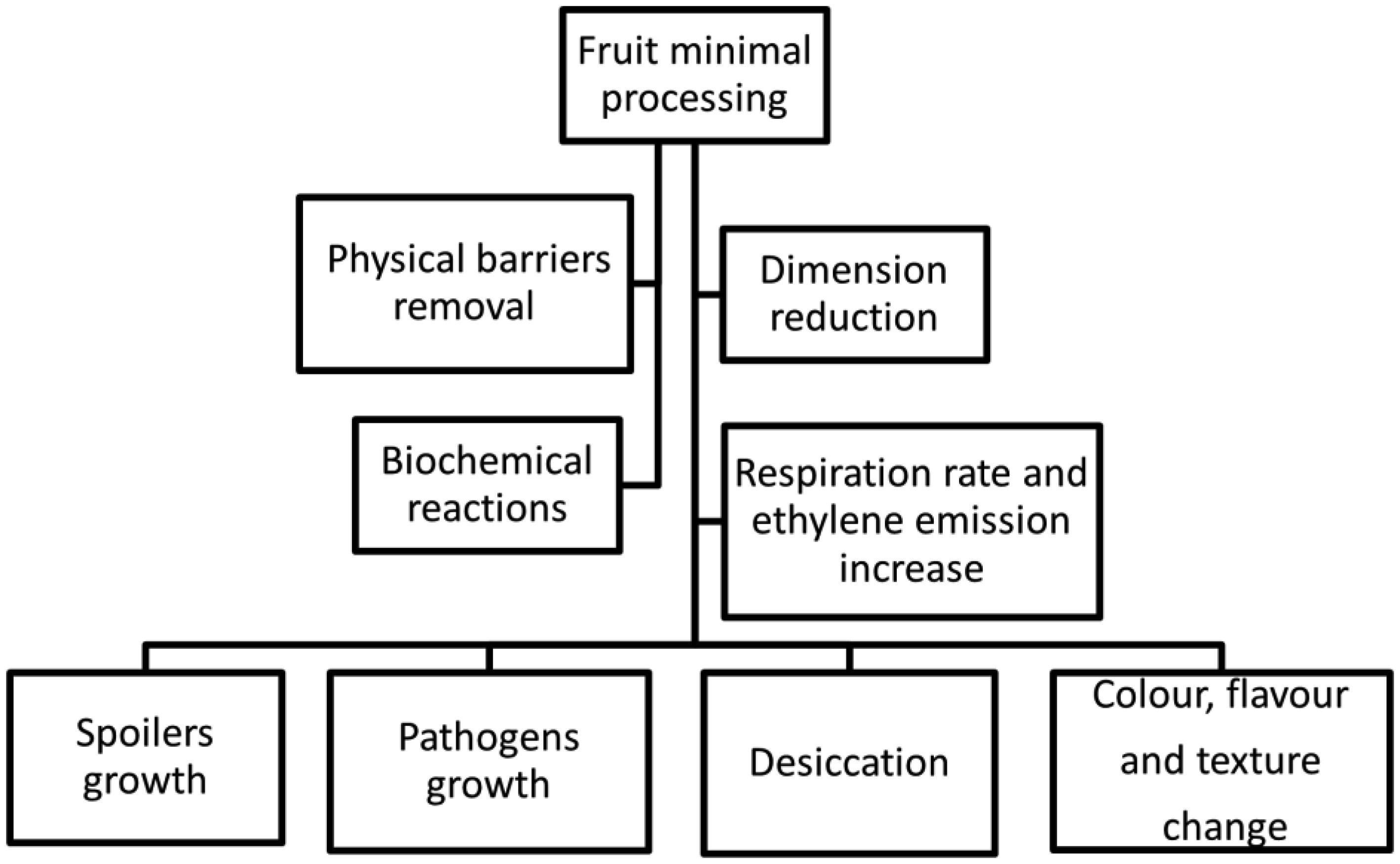
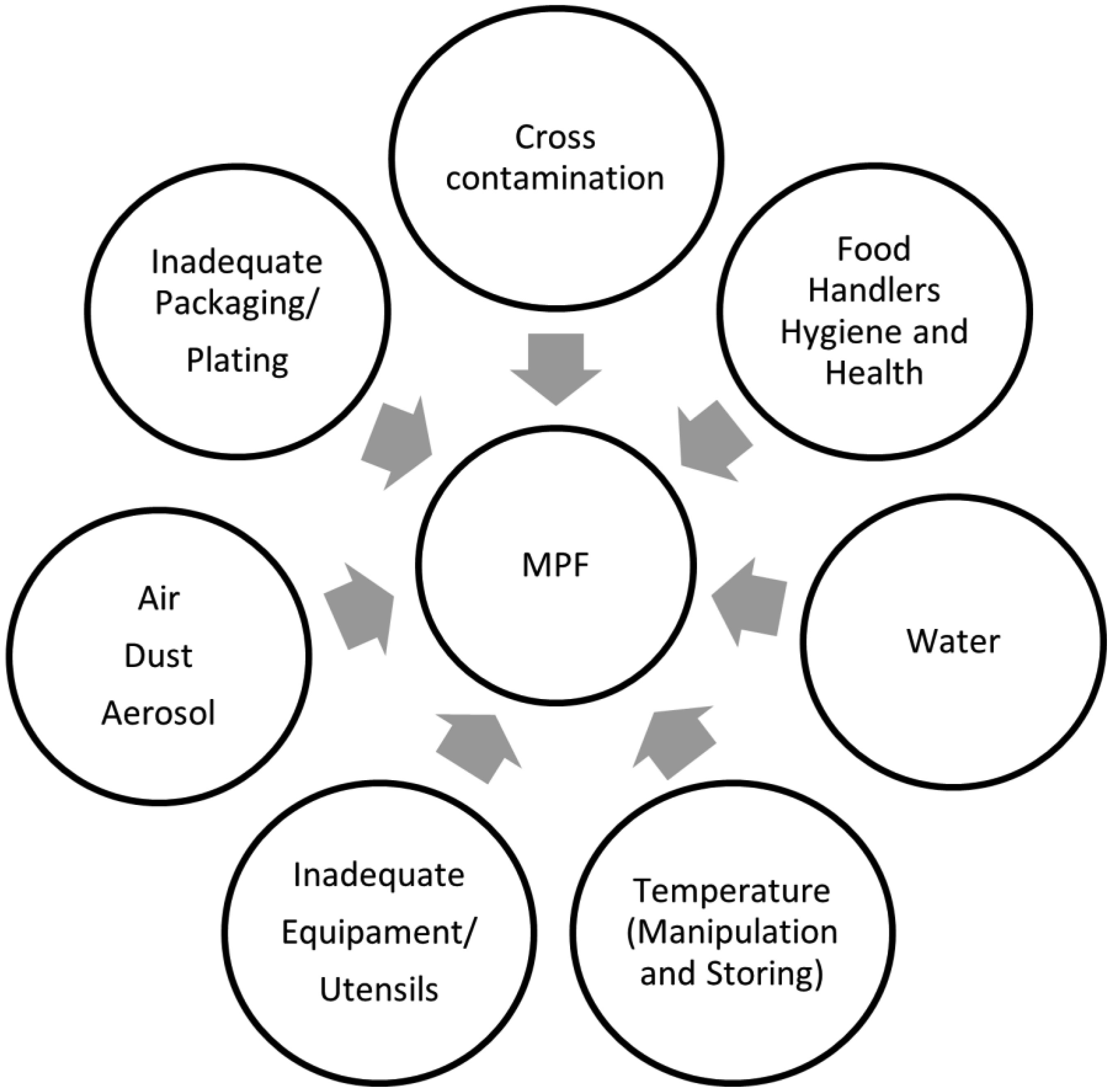
The consumption of minimally processed fruit (MPF) has increased over the last decade due to a novel trend in the food market along with the raising consumers demand for fresh, organic, convenient foods and the search for healthier lifestyles. Although represented by one of the most expanded sectors in recent years, the microbiological safety of MPF and its role as an emergent foodborne vehicle has caused great concern to the food industry and public health authorities. Such food products may expose consumers to a risk of foodborne infection as they are not subjected to prior microbial lethal methods to ensure the removal or destruction of pathogens before consumption. A considerable number of foodborne disease cases linked to MPF have been reported and pathogenic strains of Salmonella enterica, Escherichia coli, Listeria monocytogenes, as well as Norovirus accounted for the majority of cases. Microbial spoilage is also an issue of concern as it may result in huge economic losses among the various stakeholders involved in the manufacturing and commercialization of MPF. Contamination can take place at any step of production/manufacturing and identifying the nature and sources of microbial growth in the farm-to-fork chain is crucial to ensure appropriate handling practices for producers, retailers, and consumers. This review aims to summarize information about the microbiological hazards associated with the consumption of MPF and also highlight the importance of establishing effective control measures and developing coordinated strategies in order to enhance their safety.
Citation: Jessie Melo, Célia Quintas. Minimally processed fruits as vehicles for foodborne pathogens[J]. AIMS Microbiology, 2023, 9(1): 1-19. doi: 10.3934/microbiol.2023001
The consumption of minimally processed fruit (MPF) has increased over the last decade due to a novel trend in the food market along with the raising consumers demand for fresh, organic, convenient foods and the search for healthier lifestyles. Although represented by one of the most expanded sectors in recent years, the microbiological safety of MPF and its role as an emergent foodborne vehicle has caused great concern to the food industry and public health authorities. Such food products may expose consumers to a risk of foodborne infection as they are not subjected to prior microbial lethal methods to ensure the removal or destruction of pathogens before consumption. A considerable number of foodborne disease cases linked to MPF have been reported and pathogenic strains of Salmonella enterica, Escherichia coli, Listeria monocytogenes, as well as Norovirus accounted for the majority of cases. Microbial spoilage is also an issue of concern as it may result in huge economic losses among the various stakeholders involved in the manufacturing and commercialization of MPF. Contamination can take place at any step of production/manufacturing and identifying the nature and sources of microbial growth in the farm-to-fork chain is crucial to ensure appropriate handling practices for producers, retailers, and consumers. This review aims to summarize information about the microbiological hazards associated with the consumption of MPF and also highlight the importance of establishing effective control measures and developing coordinated strategies in order to enhance their safety.

| [1] |
Septembre-Malaterre A, Remize F, Poucheret P (2018) Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res Int 104: 86-99. https://doi.org/10.1016/j.foodres.2017.09.031 
|
| [2] |
Lafarga T, Colás-Medá P, Abadias M, et al. (2019) Strategies to reduce microbial risk and improve quality of fresh and processed strawberries. Inn Food Sci Emer Tech 52: 197-212. https://doi.org/10.1016/j.ifset.2018.12.012
|
| [3] |
Siroli LF, Patrignani DI, Serrazanetti F, et al. (2015) Innovative strategies based on the use of bio-control agents to improve the safety, shelf-life and quality of minimally processed fruits and vegetables. Trends Food Sci Tech 46: 302-310. https://doi.org/10.1016/j.tifs.2015.04.014
|
| [4] |
Samtiya M, Aluko RE, Dhewa T, et al. (2021) Potential health benefits of food-plant derived bioactive components: An Overview. Foods 10: 839-864. https://doi.org/10.3390/foods10040839
|
| [5] |
Prakash A, Baskaran R, Paramasivam N, et al. (2018) Essential oils based nanoemulsions to improve the microbial quality of minimally processed fruits and vegetables: A review. Food Res Int 111: 509-523. https://doi.org/10.1016/j.foodres.2018.05.066
|
| [6] |
Corato H (2019) Improving the quality and shelf-life of fresh and minimally processed fruits and vegetables for a modern industry: A comprehensive critical review from the traditional technologies into the most promising advancements. Crit Rev Food Sci Nutr 60: 940-975. https://doi.org/10.1080/10408398.2018.1553025
|
| [7] |
Oliveira M, Abadias M, Usall J, et al. (2015) Application of modified atmosphere packaging as a safety approach to fresh-cut fruits and vegetables – A review. Trends Food Sci Tech 46: 13-26. https://doi.org/10.1016/j.tifs.2015.07.017
|
| [8] |
Heaton JC, Jones K (2008) Microbial contamination of fruit and vegetables and the behavior of enteropathogens in the phyllosphere: a review. J Appl Microbiol 104: 613-626. https://doi.org/10.1111/j.1365-2672.2007.03587.x
|
| [9] |
Olaimat AN, Holley RA (2012) Factors influencing the microbial safety of fresh produce: a review. Food Microbiol 32: 1-19. https://doi.org/10.1016/j.fm.2012.04.016
|
| [10] |
Strawn LK, Schneider KR, Danyluk MD (2011) Microbial safety of tropical fruits. Crit Rev Food Sci Nutr 51: 132-145. https://doi.org/10.1080/10408390903502864
|
| [11] |
Pérez-Lavalle L, Carrasco E, Valero A (2020) Strategies for microbial decontamination of fresh blueberries and derived products. Foods 9: 1558. https://doi.org/10.3390/foods9111558
|
| [12] |
McQuiston JR, Waters RJ, Dinsmore BA, et al. (2007) Molecular determination of H antigens of Salmonella by use of microsphere-based liquid array. J Clin Microbiol 49: 565-573. https://doi.org/10.1128/JCM.01323-10
|
| [13] |
Brandl MT, Amundson R (2008) Leaf age as risk factor in contamination of lettuce with Escherichia coli O157:H7 and Salmonella enterica. Appl Environ Microbiol 74: 2298-2306. https://doi.org/10.1128/AEM.02459-07
|
| [14] |
Melotto M, Panchal S, Roy D (2014) Plant innate immunity against human bacterial pathogens. Front Microbiol 5: 411. https://doi.org/10.3389/fmicb.2014.00411
|
| [15] | Wiedemann A, Virlougeux-Payant I, Chaussé A, et al. (2014) Interaction of Salmonella with animals and plants. Front Microbiol 5: 791. https://doi.org/10.3389/fmicb.2014.00791 |
| [16] |
Coburn B, Grassi GA, Finlay BB (2007) Salmonella, the host and disease: a brief review. Immun Cell Biol 85: 112-118. https://doi.org/10.1038/sj.icb.7100007
|
| [17] |
Mali-Kale P, Winfree S, Steele-Mortimer O (2012) The bimodal lifestyle of Salmonella in epithelial cells: replication in cytosol obscures defects in vacuolar replication. PLoS One 7: e38732. https://doi.org/10.1371/journal.pone.0038732
|
| [18] | European Commission Regulation.Commission Regulation (EC) No 2073/2005 on microbiological criteria for foodstuffs. Off J Eur Union L (2005) 338: 1-26. |
| [19] |
Strawn LK, Danyluk MD (2010) Fate of Escherichia coli O157:H7 and Salmonella spp. on fresh and frozen cut mangoes and papayas. Int J Food Microbiol 138: 78-84. https://doi.org/10.1016/j.ijfoodmicro.2009.12.002
|
| [20] |
Alegre I, Abadias M, Anguera M, et al. (2010) Fate of Escherichia coli O157:H7, Salmonella and Listeria innocua on minimally-processed peaches under different storage conditions. Food Microbiol 27: 862-868. https://doi.org/10.1016/j.fm.2010.05.008
|
| [21] |
Ukuku DO, Huang L, Sommers C (2015) Efficacy of sanitizer treatments on survival and growth parameters of Escherichia coli O157:H7, Salmonella, and Listeria monocytogenes on fresh-cut pieces of Cantaloupe during storage. J Food Prot 78: 1288-1295. https://doi.org/10.4315/0362-028X.JFP-14-233
|
| [22] |
Palekar MP, Taylor TM, Maxim JE, et al. (2015) Reduction of Salmonella enterica serotype Poona and background microbiota on fresh-cut cantaloupe by electron beam irradiation. Int J Food Microbiol 202: 66-72. https://doi.org/10.1016/j.ijfoodmicro.2015.02.001
|
| [23] |
Graça A, Santo D, Quintas C, et al. (2017) Growth of Escherichia coli, Salmonella enterica and Listeria spp., and their inactivation using ultraviolet energy and electrolyzed water, on ‘Rocha’ fresh-cut pears. Food Control 77: 41-49. https://doi.org/10.1016/j.foodcont.2017.01.017
|
| [24] |
Yoo BK, Liu Y, Juneja V, et al. (2015) Growth characteristics of Shiga toxin-producing Escherichia coli (STEC) stressed by chlorine, sodium chloride, acid and starvation on lettuce and cantaloupe. Food Control 55: 97-102. https://doi.org/10.1016/j.foodcont.2015.02.040
|
| [25] |
Berger CN, Sodha SV, Shaw RK, et al. (2010) Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ Microbiol 12: 2385-2397. https://doi.org/10.1111/j.1462-2920.2010.02297.x
|
| [26] |
Croxen MA, Law RJ, Scholz R, et al. (2013) Recent advances in understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev 26: 822-880. https://doi.org/10.1128/CMR.00022-13
|
| [27] |
Solomon EB, Yaron S, Matthews KR (2002) Transmission of Escherichia coli O157:H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl Environ Microbiol 68: 397-400. https://doi.org/10.1128/AEM.68.1.397-400.2002
|
| [28] |
Natvig EE, Ingham SC, Ingham BH, et al. (2002) Salmonella enterica serovar Typhimurium and Escherichia coli contamination of root and leaf vegetables grown in soils with incorporated bovine manure. Appl Environ Microbiol 68: 2737-2744. https://doi.org/10.1128/AEM.68.6.2737-2744.2002
|
| [29] |
Abadias M, Alegre I, Oliveira M, et al. (2012) Growth potential of Escherichia coli on fresh-cut fruits (melon and pineapple) and vegetables (carrot and escarole) stored under different conditions. Food Control 27: 37-44. https://doi.org/10.1016/j.foodcont.2012.02.032
|
| [30] |
Buchanan RL, Gorris LGM, Heyman MM, et al. (2017) A review of Listeria monocytogenes: An update on outbreaks, virulence, dose response, ecology, and risk assessments. Food Control 75: 1-13. https://doi.org/10.1016/j.foodcont.2016.12.016
|
| [31] |
NicAogáin K, O'Byrne CP (2016) The role of stress and stress adaptations determining the fate of bacterial pathogen Listeria monocytogenes in the food chain. Front Microbiol 7: 1865. https://doi.org/10.3389/fmicb.2016.01865
|
| [32] |
Ferreira V, Wiedmann M, Teixeira P, et al. (2014) Listeria monocytogenes persistence in food-associated environments: Epidemiology, strain characteristics, and implications for public health. J Food Prot 77: 150-170. https://doi.org/10.4315/0362-028X.JFP-13-150
|
| [33] |
Colagiorgi A, Bruini I, Di Ciccio PA, et al. (2017) Listeria monocytogenes biofilms in the wonderland of food industry. Pathogens 6: 41. https://doi.org/10.3390/pathogens6030041
|
| [34] |
Colás Medà P, Vinas I, Alegre I, et al. (2017) The impact of a cold chain break on the survival of Salmonella enterica and Listeria monocytogenes on minimally processed “Conference” pears during their shelf life. J Sci Food Agric 97: 3077-3080. https://doi.org/10.1002/jsfa.8127
|
| [35] |
Melo J, Andrew PW, Faleiro ML (2015) Listeria monocytogenes in cheese and the dairy environment remains a food safety challenge: The role of stress responses. Food Res Int 67: 75-90. https://doi.org/10.1016/j.foodres.2014.10.031
|
| [36] | Shamloo E, Hosseini H, Moghadam ZA, et al. (2019) Importance of Listeria monocytogenes in food safety: a review of its prevalence, detection, and antibiotic resistance. Iran J Vet Res 20: 241-254. |
| [37] |
Garner D, Kathariou S (2016) Fresh produce-associated listeriosis outbreaks, sources of concern, teachable moments, and insights. J Food Prot 79: 337-344. https://doi.org/10.4315/0362-028X.JFP-15-387
|
| [38] |
Lopez-Valladares G, Danielsson-Tham ML, Tham W (2018) Implicated food products for listeriosis and changes in serovars of Listeria monocytogenes affecting humans for recent decades. Foodborne Pathog Dis 15: 387-397. https://doi.org/10.1089/fpd.2017.2419
|
| [39] |
Gustafson RE, Ryser ET (2017) Thermal inactivation and growth of Listeria monocytogenes during production and storage of caramel apples. Food Control 79: 234-238. https://doi.org/10.1016/j.foodcont.2017.03.043
|
| [40] | Glass KA, Golden MC, Wanless BJ, et al. (2015) Growth of Listeria monocytogenes within a caramel-coated apple microenvironment. AMS mBio 6: e01232-15. https://doi.org/10.1128/mBio.01232-15 |
| [41] |
Ruiz-Llacsahuanga B, Hamilton A, Zaches R, et al. (2016) Utility of rapid tests to assess the prevalence of indicator organisms (aerobic plate count, Enterobacteriaceae, coliforms, Escherichia coli, and Listeria spp.) in apple packinghouses. Int J Food Microbiol 337: 108949. https://doi.org/10.1016/j.ijfoodmicro.2020.108949
|
| [42] |
De Jesus AJ, Sheth I, Kwon HJ, et al. (2020) Survival of a serotype 4b strain and a serotype 1/2a strain of Listeria monocytogenes, isolated from a stone fruit outbreak investigation, on whole stone fruit at 4 °C. Int J Food Microbiol 334: 108801. https://doi.org/10.1016/j.ijfoodmicro.2020.108801
|
| [43] |
Conway WS, Leverentz B, Saftner RA, et al. (2000) Survival and growth of Listeria monocytogenes on fresh-cut apples slices and its interactions with Glomerella cingulata and Penicillium expansum. Plant Dis 84: 177-181. https://doi.org/10.1094/PDIS.2000.84.2.177
|
| [44] | Ziegler M, Rüegg S, Stephan R, et al. (2018) Growth potential of Listeria monocytogenes in six different RTE fruit products: impact of food matrix, storage temperature and shelf life. Ital J Food Saf 7: 7581. https://doi.org/10.4081/ijfs.2018.7581 |
| [45] |
Carpentier B, Cerf O (2011) Review–persistence of Listeria monocytogenes in food industry equipment and premises. Int J Food Microbiol 145: 1-8. https://doi.org/10.1016/j.ijfoodmicro.2011.01.005
|
| [46] |
Bosch A, Gkogka E, Le Guyader FS, et al. (2018) Foodborne viruses: Detection, risk assessment, and control options in food processing. Int J Food Microbiol 285: 110-128. https://doi.org/10.1016/j.ijfoodmicro.2018.06.001
|
| [47] |
Marsh Z, Shah MP, Wikswo ME (2018) Epidemiology of foodborne norovirus outbreaks -United States, 2009–2015. Food Saf (Tokyo) 6: 58-66. https://doi.org/10.14252/foodsafetyfscj.2017028
|
| [48] |
Butot S, Putallaz T, Sánchez G (2008) Effects of sanitation, freezing and frozen storage on enteric viruses in berries and herbs. Int J Food Microbiol 126: 30-5. https://doi.org/10.1016/j.ijfoodmicro.2008.04.033
|
| [49] |
Le Guyader FS, Mittelholzer C, Haugarreau L, et al. (2004) Detection of noroviruses in raspberries associated with a gastroenteritis outbreak. Int J Food Microbiol 97: 179-186. https://doi.org/10.1016/j.ijfoodmicro.2004.04.018
|
| [50] |
Quansah JK, Gazula H, Holland R, et al. (2019) Microbial quality of blueberries for the fresh market. Food Control 100: 92-96. https://doi.org/10.1016/j.foodcont.2018.12.034
|
| [51] |
Bernard H, Faber M, Wilking H, et al. (2014) Large multistate outbreak of norovirus gastroenteritis associated with frozen strawberries, Germany, 2012. Euro Surveill 19: 20719. https://doi.org/10.2807/1560-7917.ES2014.19.8.20719
|
| [52] |
Sarvikivi E, Roivainen M, Maunula L, et al. (2012) Multiple norovirus outbreaks linked to imported frozen raspberries. Epidemiol Infect 140: 260-267. https://doi.org/10.1017/S0950268811000379
|
| [53] |
Warriner K, Ibrahim F, Dickinson M, et al. (2013) Internalization of human pathogens within growing salad vegetables. Biotechnol Genet Eng Rev 20: 117-136. https://doi.org/10.1080/02648725.2003.10648040
|
| [54] |
Lopman B, Gastañaduy P, Park GW, et al. (2012) Environmental transmission of norovirus gastroenteritis. Curr Opin Virol 2: 96-102. https://doi.org/10.1016/j.coviro.2011.11.005
|
| [55] |
Harris LJ, Farber JN, Beuchat LR, et al. (2003) Outbreaks associated with fresh produce: incidence, growth, and survival of pathogens in fresh and fresh-cut produce. Compr Rev Food Sci Food Saf 2: 78-141. https://doi.org/10.1111/j.1541-4337.2003.tb00031.x
|
| [56] |
Alegbeleye OO, Singleton I, Sant'Ana AS (2018) Sources and contamination routes of microbial pathogens to fresh produce during field cultivation: A review. Food Microbiol 73: 177-208. https://doi.org/10.1016/j.fm.2018.01.003
|
| [57] |
Chatziprodromidou IP, Bellou G, Vantarakis G, et al. (2018) Viral outbreaks linked to fresh produce consumption: a systematic review. J Appl Microbiol 124: 932-942. https://doi.org/10.1111/jam.13747
|
| [58] |
Vasickova P, Pavlik I, Verani M, et al. (2010) Issues concerning survival of viruses on surfaces. Food Environ Virol 2: 24-34. https://doi.org/10.1007/s12560-010-9025-6
|
| [59] |
Teunis PFM, Le Guyader FS, Liu P (2020) Noroviruses are highly infectious but there is strong variation in host susceptibility and virus pathogenicity. Epidemics 32: 100401. https://doi.org/10.1016/j.epidem.2020.100401
|
| [60] |
Baert L, Debevere J, Uyttendaele M (2009) The efficacy of preservation methods to inactivate foodborne viruses. Int J Food Microbiol 131: 83-94. https://doi.org/10.1016/j.ijfoodmicro.2009.03.007
|
| [61] |
Terio V, Bottaro M, Pavoni E, et al. (2017) Occurrence of hepatitis A and E and norovirus GI and GII in ready-to-eat vegetables in Italy. Int J Food Microbiol 249: 61-65. https://doi.org/10.1016/j.ijfoodmicro.2017.03.008
|
| [62] |
D'Souza D, Sair A, Williams K, et al. (2006) Persistence of caliciviruses on environmental surfaces and their transfer to food. Int J Food Microbiol 108: 84-91. https://doi.org/10.1016/j.ijfoodmicro.2005.10.024
|
| [63] |
Mattison K, Karthikeyan K, Abebe M, et al. (2007) Survival of calicivirus in foods and on surfaces: experiments with feline calicivirus as a surrogate for norovirus. J Food Prot 70: 500-503. https://doi.org/10.4315/0362-028X-70.2.500
|
| [64] |
Escudero-Abarca B, Rawsthorne H, Gensel C, et al. (2012) Persistence and transferability of Noroviruses on and between common surfaces and foods. J Food Prot 75: 927-35. https://doi.org/10.4315/0362-028x.jfp-11-460
|
| [65] | Gutierrez-Rodrigues E, Adhikari A (2018) Pre-harvesting farming practices impacting quality of fresh produce safety. ASM Microbiol Spectrum 6: 1-20. https://doi.org/10.1128/microbiolspec.PFS-0022-2018 |
| [66] |
Yeni F, Alpas H (2017) Vulnerability of global food production to extreme climatic events. Food Res Int 96: 27-39. https://doi.org/10.1016/j.foodres.2017.03.020
|
| [67] |
Machado-Moreira B, Richards K, Brennan F, et al. (2019) Microbial contamination of fresh produce: What, where and how?. Compr Rev Food Sci Food Saf 18: 1727-1750. https://doi.org/10.1111/1541-4337.12487
|
| [68] |
Gil MI, Selma MV, López-Gálvez F, et al. (2009) Fresh cut product sanitation and wash water disinfection: Problems and solutions. Int J Food Microbiol 134: 37-45. https://doi.org/10.1016/j.ijfoodmicro.2009.05.021
|
| [69] |
Marín A, Tudela JA, Garrido Y, et al. (2020) Chlorinated wash water and pH regulators affect chlorine gas emission and disinfection byproducts. Innov Food Sci Emerg Technol 66: 102533. https://doi.org/10.1016/j.ifset.2020.102533
|
| [70] |
Penteado AL, Castro MFPM, Rezende ACB (2014) Salmonella enterica serovar Enteritidis and Listeria monocytogenes in mango (Mangifera indica L.) pulp: growth, survival and cross-contamination. J Sci Food Agric 99: 2746-2751. https://doi.org/10.1002/jsfa.6619
|
| [71] |
Lehto M, Kuisma R, Maatta J, et al. (2011) Hygienic level and surface contamination in fresh-cut vegetable production plants. Food Control 22: 469-475. https://doi.org/10.1016/j.foodcont.2010.09.029
|
| [72] |
Cunault C, Faille C, Calabozo-Delgado A, et al. (2019) Structure and resistance to mechanical stress and enzymatic cleaning of Pseudomonas fluorescens biofilms formed in fresh-cut ready to eat washing tanks. J Food Eng 262: 154-161. https://doi.org/10.1016/j.jfoodeng.2019.06.006
|
| [73] |
McCollum JT, Cronquist AB, Silk BJ, et al. (2013) Multistate outbreak of listeriosis associated with cantaloupe. N J Engl J Med 369: 944-953. https://doi.org/10.1056/NEJMoa1215837
|
| [74] |
Corbo MR, Campaniello D, D'Amato D, et al. (2005) Behaviour of Listeria monocytogenes and Escherichia coli O157:H7 in fresh-sliced cactus-pear fruit. J Food Safe 25: 157-172. https://doi.org/10.1111/j.1745-4565.2005.00570.x
|
| [75] | Raybaudi-Massilia RM, Mosqueda-Melgar J, Sobrino-Lopez A, et al. (2009) Use of malic acid and other quality stabilizing compounds to assure the safety of fresh-cut “Fuji” apples by inactivation of Listeria monocytogenes, Salmonella Enteritidis and Escherichia coli O157:H7. J Food Safe 29: 239-252. https://doi.org/10.1111/j.1745-4565.2009.00153.x |
| [76] |
Erickson MC (2012) Internalization of fresh produce by foodborne pathogens. Annu Rev Food Sci Technol 3: 283-310. https://doi.org/10.1146/annurev-food-022811-101211
|
| [77] |
Strobel G (2018) The emergence of endophytic microbes and their biological promise. J Fungi 4: 57-76. https://doi.org/10.3390/jof4020057
|
| [78] |
Gibbs DS, Anderson GL, Beuchat LR, et al. (2005) Potential role of Diploscapter sp. strain LKC25, a bacterivorous nematode from soil, as a vector of food-borne pathogenic bacteria to preharvest fruits and vegetables. Appl Environ Microbiol 71: 2433-2437. https://doi.org/10.1128/AEM.71.5.2433-2437.2005
|
| [79] |
Lim JA, Lee DH, Heu S (2014) The interaction of human enteric pathogens with plants. Plant Pathol 30: 109-116. https://doi.org/10.5423/PPJ.RW.04.2014.0036
|
| [80] |
Cools D, Merckx R, Vlassak K, et al. (2001) Survival of E. coli and Enterococcus derived from pig slurry in soils with different texture. Appl Soil Ecol 17: 53-62. https://doi.org/10.1016/S0929-1393(00)00133-5
|
| [81] |
López-Gálvez F, Gil MI, Truchado P, et al. (2010) Cross-contamination of fresh-cut lettuce after a short-term exposure during pre-washing cannot be controlled after subsequent washing with chlorine dioxide or sodium hypochlorite. Food Microbiol 27: 199-204. https://doi.org/10.1016/j.fm.2009.09.009
|
| [82] |
Holden N, Pritchard L, Toth I (2009) Colonization out with the colon: plants as an alternative environmental reservoir for human pathogenic enterobacteria. FEMS Microbiol Rev 33: 689-703. https://doi.org/10.1111/j.1574-6976.2008.00153.x
|
| [83] |
Deering AJ, Mauer LJR, Pruitt RE (2012) Internalization of E. coli O157:H7 and Salmonella spp. in plants: A review. Food Res Int 45: 567-575. https://doi.org/10.1016/j.foodres.2011.06.058
|
| [84] |
Gautam D, Dobhal S, Peyton ME, et al. (2014) Surface survival and internalization of Salmonella through natural cracks on developing cantaloupe fruits alone or in the presence of the melon wilt Erwinia tracheiphila. PLoS One 9: e105248. https://doi.org/10.1371/journal.pone.0105248
|
| [85] |
Chen LQ, Hou B, Lalonde H, et al. (2010) Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 468: 527-532. https://doi.org/10.1038/nature09606
|
| [86] |
Martínez-Vaz BM, Fink RC, Diez-Gonzalez F, et al. (2014) Enteric pathogen-plant interactions: molecular connections leading to colonization and growth and implications for foin od safety. Microbes Environ 29: 123-135. https://doi.org/10.1264/jsme2.ME13139
|
| [87] | Fatima U, Senthil-Kumar M (2015) Plant and pathogen nutrient acquisition strategies. Front Plant Scien 6: 1-12. https://doi.org/10.3389/fpls.2015.00750 |
| [88] |
Oms-Oliu G, Rojas-Grau MA, Gonzalez LA, et al. (2010) Recent approaches using chemical treatments to preserve quality of fresh-cut fruit: A review. Postharvest Biol Tech 57: 139-148. https://doi.org/10.1016/j.postharvbio.2010.04.001
|
| [89] |
Luo Y, Nou X, Yang Y, et al. (2011) Determination of free chlorine concentrations needed to prevent Escherichia coli O157:H7 cross-contamination during fresh cut produce wash. J Food Prot 24: 352-358. https://doi.org/10.4315/0362-028X.JFP-10-429
|
| [90] |
Beuchat LR (2002) Ecological factors influencing survival and growth of human pathogens on raw fruits and vegetables. Microbes Infect 4: 413-423. https://doi.org/10.1016/S1286-4579(02)01555-1
|
Figures(2) / Tables(2)

Jessie Melo, Célia Quintas. Minimally processed fruits as vehicles for foodborne pathogens[J]. AIMS Microbiology, 2023, 9(1): 1-19. doi: 10.3934/microbiol.2023001







 DownLoad:
DownLoad: