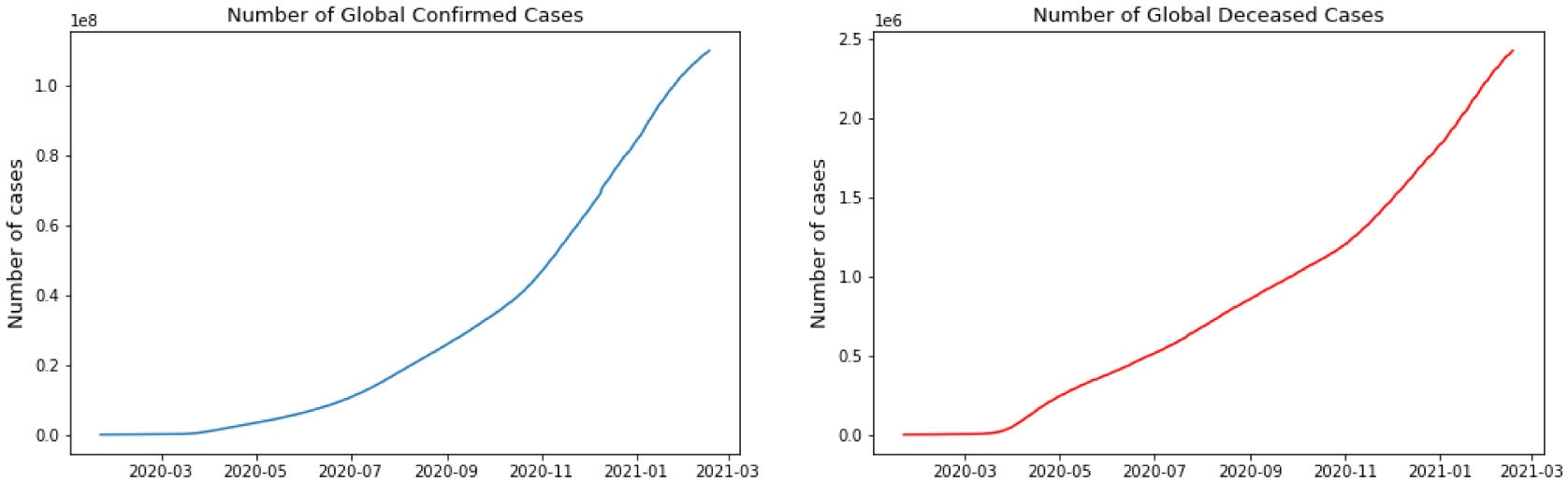
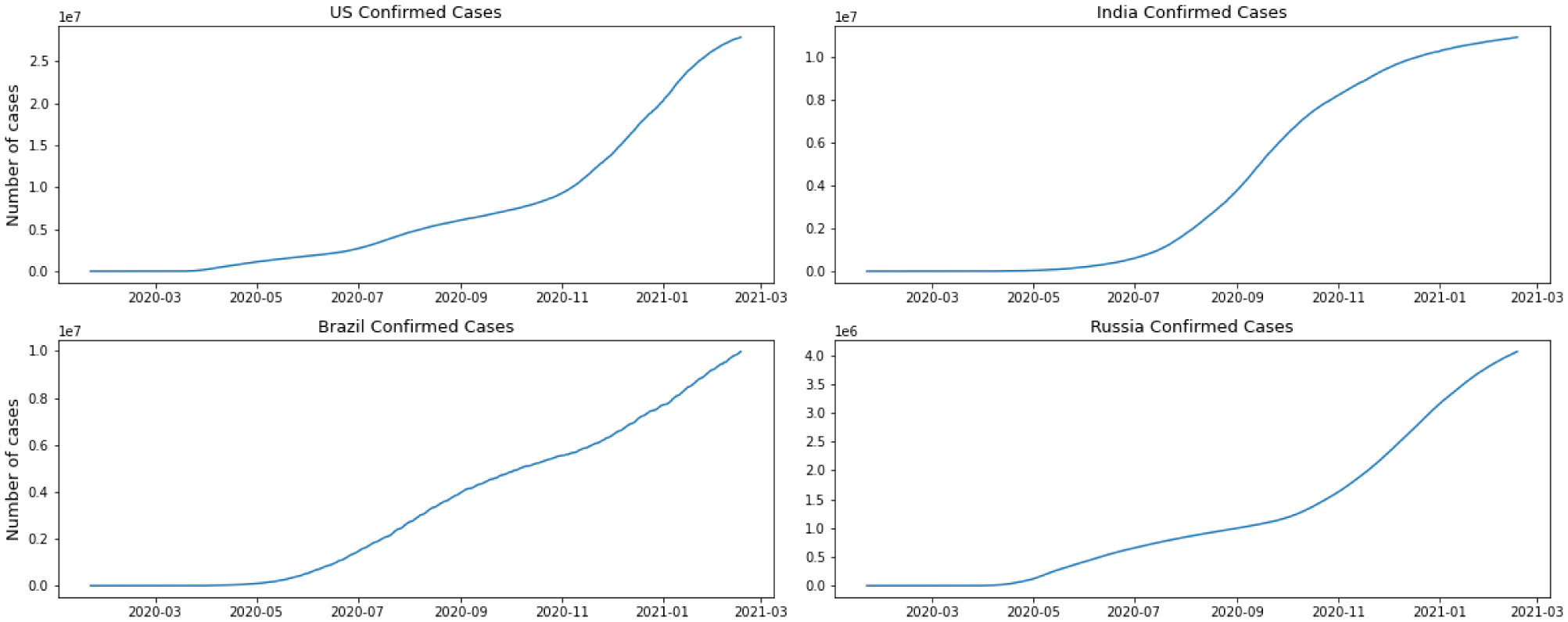
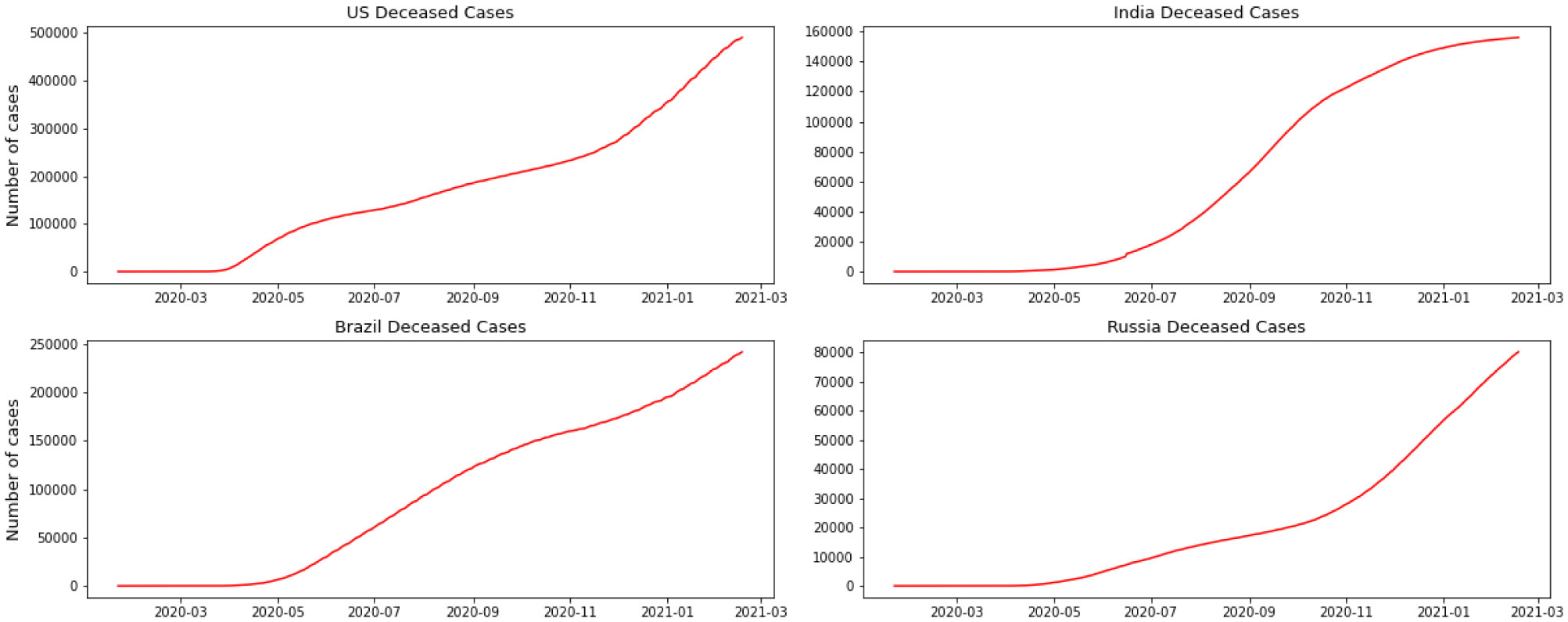
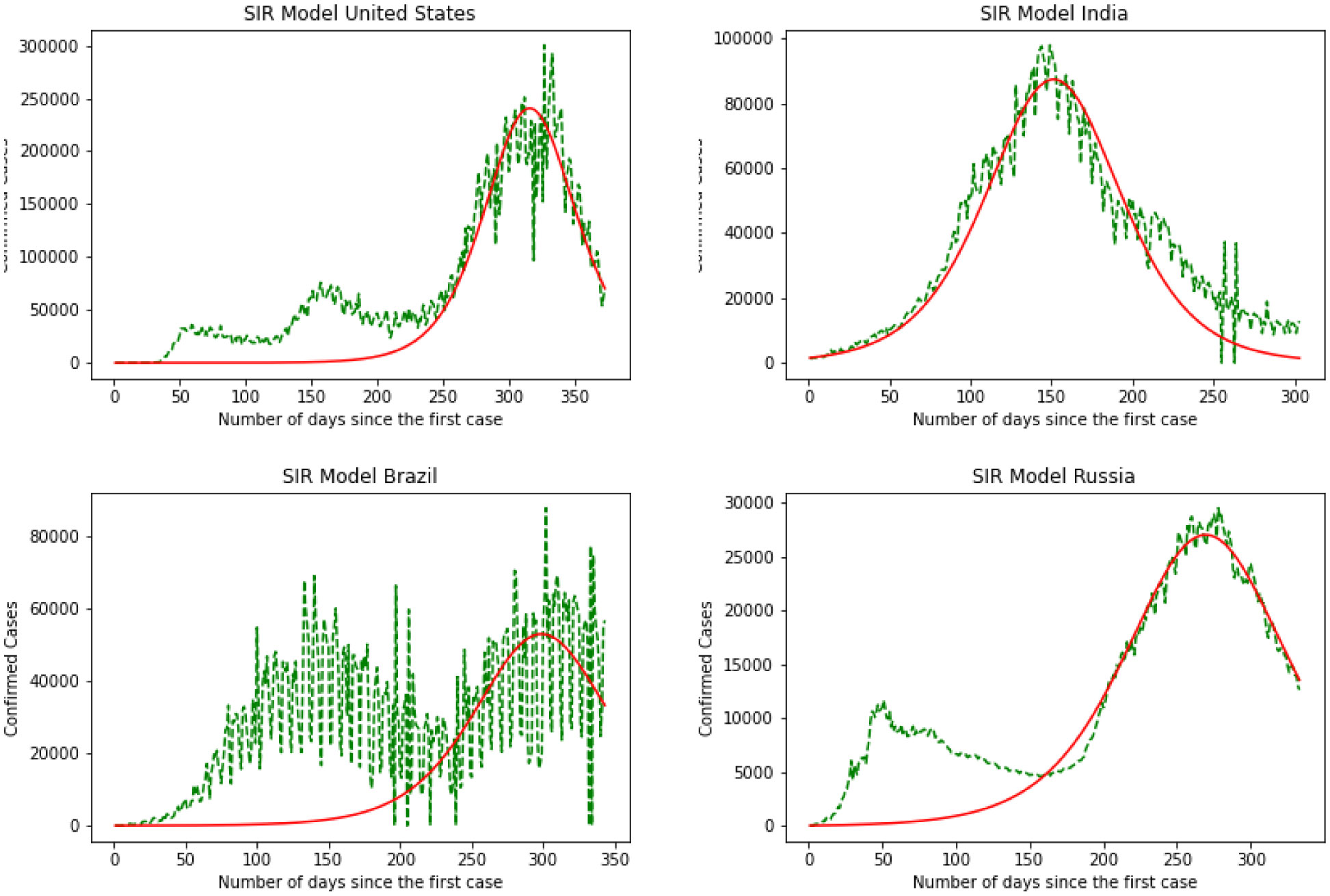
Starting February 2020, COVID-19 was confirmed in 11,946 people worldwide, with a mortality rate of almost 2%. A significant number of epidemic diseases consisting of human Coronavirus display patterns. In this study, with the benefit of data analytic, we develop regression models and a Susceptible-Infected-Recovered (SIR) model for the contagion to compare the performance of models to predict the number of cases. First, we implement a good understanding of data and perform Exploratory Data Analysis (EDA). Then, we derive parameters of the model from the available data corresponding to the top 4 regions based on the history of infections and the most infected people as of the end of August 2020. Then models are compared, and we recommend further research.
Citation: Saina Abolmaali, Samira Shirzaei. A comparative study of SIR Model, Linear Regression, Logistic Function and ARIMA Model for forecasting COVID-19 cases[J]. AIMS Public Health, 2021, 8(4): 598-613. doi: 10.3934/publichealth.2021048
Starting February 2020, COVID-19 was confirmed in 11,946 people worldwide, with a mortality rate of almost 2%. A significant number of epidemic diseases consisting of human Coronavirus display patterns. In this study, with the benefit of data analytic, we develop regression models and a Susceptible-Infected-Recovered (SIR) model for the contagion to compare the performance of models to predict the number of cases. First, we implement a good understanding of data and perform Exploratory Data Analysis (EDA). Then, we derive parameters of the model from the available data corresponding to the top 4 regions based on the history of infections and the most infected people as of the end of August 2020. Then models are compared, and we recommend further research.

| [1] |
Porta M (2014) A dictionary of epidemiology Oxford university press. doi: 10.1093/acref/9780199976720.001.0001

|
| [2] | WHO COVID-19 Epidemic disease Available from: https://www.who.int/emergencies/diseases/news. |
| [3] | AJMC Staff A Timeline of COVID-19 Developments in 2020 (2021) .Available from: https://www.ajmc.com/view/a-timeline-of-covid19-developments-in-2020. |
| [4] | COVID-19 CORONAVIRUS PANDEMIC Available from: https://www.worldometers.info/coronavirus/. |
| [5] |
Bontempi E, Coccia M (2021) International trade as critical parameter of COVID-19 spread that outclasses demographic, economic, environmental, and pollution factors. Environ Res 201: 111514. doi: 10.1016/j.envres.2021.111514
|
| [6] |
Bontempi E (2020) Commercial exchanges instead of air pollution as possible origin of COVID-19 initial diffusion phase in Italy: more efforts are necessary to address interdisciplinary research. Environ Res 188: 109775. doi: 10.1016/j.envres.2020.109775
|
| [7] |
Bontempi E, Coccia M, Vergalli S, et al. (2021) Can commercial trade represent the main indicator of the COVID-19 diffusion due to human-to-human interactions? A comparative analysis between Italy, France, and Spain. Environ Res 201: 111529. doi: 10.1016/j.envres.2021.111529
|
| [8] |
Anand U, Cabreros C, Mal J, et al. (2021) Novel coronavirus disease 2019 (COVID-19) pandemic: From transmission to control with an interdisciplinary vision. Environ Res 197: 111126. doi: 10.1016/j.envres.2021.111126
|
| [9] |
Bontempi E, Vergalli S, Squazzoni F (2020) Understanding COVID-19 diffusion requires an interdisciplinary, multi-dimensional approach. Environ Res 188: 109814. doi: 10.1016/j.envres.2020.109814
|
| [10] |
Al Huraimel K, Alhosani M, Kunhabdulla S, et al. (2020) SARS-CoV-2 in the environment: Modes of transmission, early detection and potential role of pollutions. Sci Total Environ 744: 140946. doi: 10.1016/j.scitotenv.2020.140946
|
| [11] |
Yuan J, Li M, Lv G, et al. (2020) Monitoring transmissibility and mortality of COVID-19 in Europe. Int J Infect Dis 95: 311-315. doi: 10.1016/j.ijid.2020.03.050
|
| [12] |
Liu Y, Gayle A, Wilder-Smith A, et al. (2020) The reproductive number of COVID-19 is higher compared to SARS coronavirus. J Travel Med 27: taaa021. doi: 10.1093/jtm/taaa021
|
| [13] |
Rosario D, Mutz Y, Bernardes P, et al. (2020) Relationship between COVID-19 and weather: Case study in a tropical country. Int J Hyg Environ Health 229: 113587. doi: 10.1016/j.ijheh.2020.113587
|
| [14] |
Coccia M (2020) Factors determining the diffusion of COVID-19 and suggested strategy to prevent future accelerated viral infectivity similar to COVID. Sci Total Environ 729: 138474. doi: 10.1016/j.scitotenv.2020.138474
|
| [15] |
Coccia M (2021) The effects of atmospheric stability with low wind speed and of air pollution on the accelerated transmission dynamics of COVID-19. Int J Environ Stud 78: 1-27. doi: 10.1080/00207233.2020.1802937
|
| [16] |
Coccia M (2021) High health expenditures and low exposure of population to air pollution as critical factors that can reduce fatality rate in COVID-19 pandemic crisis: a global analysis. Environ Res 199: 111339. doi: 10.1016/j.envres.2021.111339
|
| [17] |
Coccia M (2021) Effects of the spread of COVID-19 on public health of polluted cities: results of the first wave for explaining the dej vu in the second wave of COVID-19 pandemic and epidemics of future vital agents. Environ Sci Pollut Res Int 28: 19147-19154. doi: 10.1007/s11356-020-11662-7
|
| [18] |
Coccia M (2021) How do low wind speeds and high levels of air pollution support the spread of COVID-19? Atmos Pollut Res 12: 437-445. doi: 10.1016/j.apr.2020.10.002
|
| [19] |
Coccia M (2020) An index to quantify environmental risk of exposure to future epidemics of the COVID-19 and similar viral agents: Theory and practice. Environ Res 191: 110155. doi: 10.1016/j.envres.2020.110155
|
| [20] | Coccia M (2021) Preparedness of countries to face covid-19 pandemic crisis: Strategic positioning and underlying structural factors to support strategies of prevention of pandemic threats. Environ Res 111678. |
| [21] |
Coccia M (2021) The relation between length of lockdown, numbers of infected people and deaths of Covid-19, and economic growth of countries: Lessons learned to cope with future pandemics similar to Covid-19 and to constrain the deterioration of economic system. Sci Total Environ 775: 145801. doi: 10.1016/j.scitotenv.2021.145801
|
| [22] |
Bashir M, Jiang B, Komal B, et al. (2020) Correlation between environmental pollution indicators and COVID-19 pandemic: a brief study in Californian context. Environ Res 187: 109652. doi: 10.1016/j.envres.2020.109652
|
| [23] | Abolmaali S, Roodposhti F (2018) Portfolio Optimization Using Ant Colony Method a Case Study on Tehran Stock Exchange. J Account 8. |
| [24] | Centers for Disease Control and Prevention, National Center for Immunization and Respiratory Diseases (NCIRD) 1918 Pandemic (H1N1 virus) Available from: https://www.cdc.gov/flu/pandemic-resources/1918-pandemic-h1n1.html. |
| [25] | Centers for Disease Control and Prevention, National Center for Immunization and Respiratory Diseases (NCIRD) 2009 H1N1 Pandemic (H1N1pdm09 virus) Available from: https://www.cdc.gov/flu/pandemic-resources/2009-h1n1-pandemic.html. |
| [26] | Ebola Lessons for Global Health and PPE Preparedness during Outbreak Available from: https://www.derekduck.com/page/267. |
| [27] |
Chowell G, Sattenspiel L, Bansal S, et al. (2016) Mathematical models to characterize early epidemic growth: A review. Phys Life Rev 18: 66-97. doi: 10.1016/j.plrev.2016.07.005
|
| [28] |
Mutalik A (2017) Models to predict H1N1 outbreaks: a literature review. Int J Community Med Public Health 4: 3068-3075. doi: 10.18203/2394-6040.ijcmph20173814
|
| [29] |
Zhan C, Chi K, Lai Z, et al. (2020) Prediction of COVID-19 Spreading Profiles in South Korea, Italy and Iran by Data-Driven Coding. PLoS One 15: e0234763. doi: 10.1371/journal.pone.0234763
|
| [30] | Lover A, McAndrew T (2020) Sentinel Event Surveillance to Estimate Total SARS-CoV-2 Infections, United States. MedRxiv . |
| [31] | Liu P, Beeler P, Chakrabarty R (2020) COVID-19 Progression Timeline and Effectiveness of Response-to-Spread Interventions across the United States. MedRxiv . |
| [32] | Abolmaali S, Shirzaei S (2021) Forecasting COVID-19 Number of Cases by Implementing ARIMA and SARIMA with Grid Search in the United States. MedRxiv . |
| [33] | Roosa K, Lee Y, Luo R, et al. (2020) Real-time forecasts of the COVID-19 epidemic in China from February 5th to February 24th, 2020. Infect Dis Model 5: 256-263. |
| [34] | Gupta R, Pal S (2020) Trend Analysis and Forecasting of COVID-19 outbreak in India. MedRxiv . |
| [35] |
Moein S, Nickaeen N, Roointan A, et al. (2021) Inefficiency of SIR models in forecasting COVID-19 epidemic: a case study of Isfahan. Sci Rep 11: 4725. doi: 10.1038/s41598-021-84055-6
|
| [36] |
Calafiore G, Novara C, Possieri C (2020) A modified SIR model for the COVID-19 contagion in Italy. Annu Rev Control 50: 361-372. doi: 10.1016/j.arcontrol.2020.10.005
|
| [37] |
Satsuma J, Willox R, Ramani A, et al. (2004) Extending the SIR epidemic model. Physica A: Statistical Mechanics And Its Applications 369-375. doi: 10.1016/j.physa.2003.12.035
|
| [38] |
Anastassopoulou C, Russo L, Tsakris A, et al. (2020) Data-based analysis, modelling and forecasting of the COVID-19 outbreak. PloS One 15: e0230405. doi: 10.1371/journal.pone.0230405
|
| [39] |
Read J, Bridgen J, Cummings D, et al. (2021) Novel coronavirus 2019-nCoV: early estimation of epidemiological parameters and epidemic predictions. Philos Trans R Soc Lond B Biol Sci 376: 20200265. doi: 10.1098/rstb.2020.0265
|
| [40] |
Lin Q, Zhao S, Gao D, et al. (2020) A conceptual model for the coronavirus disease 2019 (COVID-19) outbreak in Wuhan, China with individual reaction and governmental action. Int J Infect Dis 93: 211-216. doi: 10.1016/j.ijid.2020.02.058
|
| [41] |
Giordano G, Blanchini F, Bruno R, et al. (2020) Modelling the COVID-19 epidemic and implementation of population-wide interventions in Italy. Nat Med 26: 855-860. doi: 10.1038/s41591-020-0883-7
|
| [42] | Roda W, Varughese M, Han D, et al. (2020) Why is it difficult to accurately predict the COVID-19 epidemic? Infect Dis Model 5: 271-281. |
| [43] |
Furtado P (2021) Epidemiology SIR with Regression, Arima, and Prophet in Forecasting Covid-19. Eng Proc 5: 52. doi: 10.3390/engproc2021005052
|
| [44] | Abuhasel K, Khadr M, Alquraish M (2020) Analyzing and forecasting COVID-19 pandemic in the Kingdom of Saudi Arabia using ARIMA and SIR models. Comput Intell . |
| [45] |
Diekmann O, Heesterbeek J (2008) Mathematical epidemiology of infectious diseases: model building, analysis and interpretation. Math Biosci 213: 1-12. doi: 10.1016/j.mbs.2008.02.005
|
| [46] | Kermack W, McKendrick A (1991) Contributions to the mathematical theory of epidemics. Bull Math Biol 53: 33-55. |
| [47] |
Bjrnstad O, Finkenstdt B, Grenfell B (2002) Dynamics of measles epidemics: estimating scaling of transmission rates using a time series SIR model. Ecol Monogr 72: 169-184. doi: 10.1890/0012-9615(2002)072[0169:DOMEES]2.0.CO;2
|
| [48] | Tabachnick BG, Fidell LS, Ullman JB (2007) Using multivariate statistics Boston, MA: Pearson. |

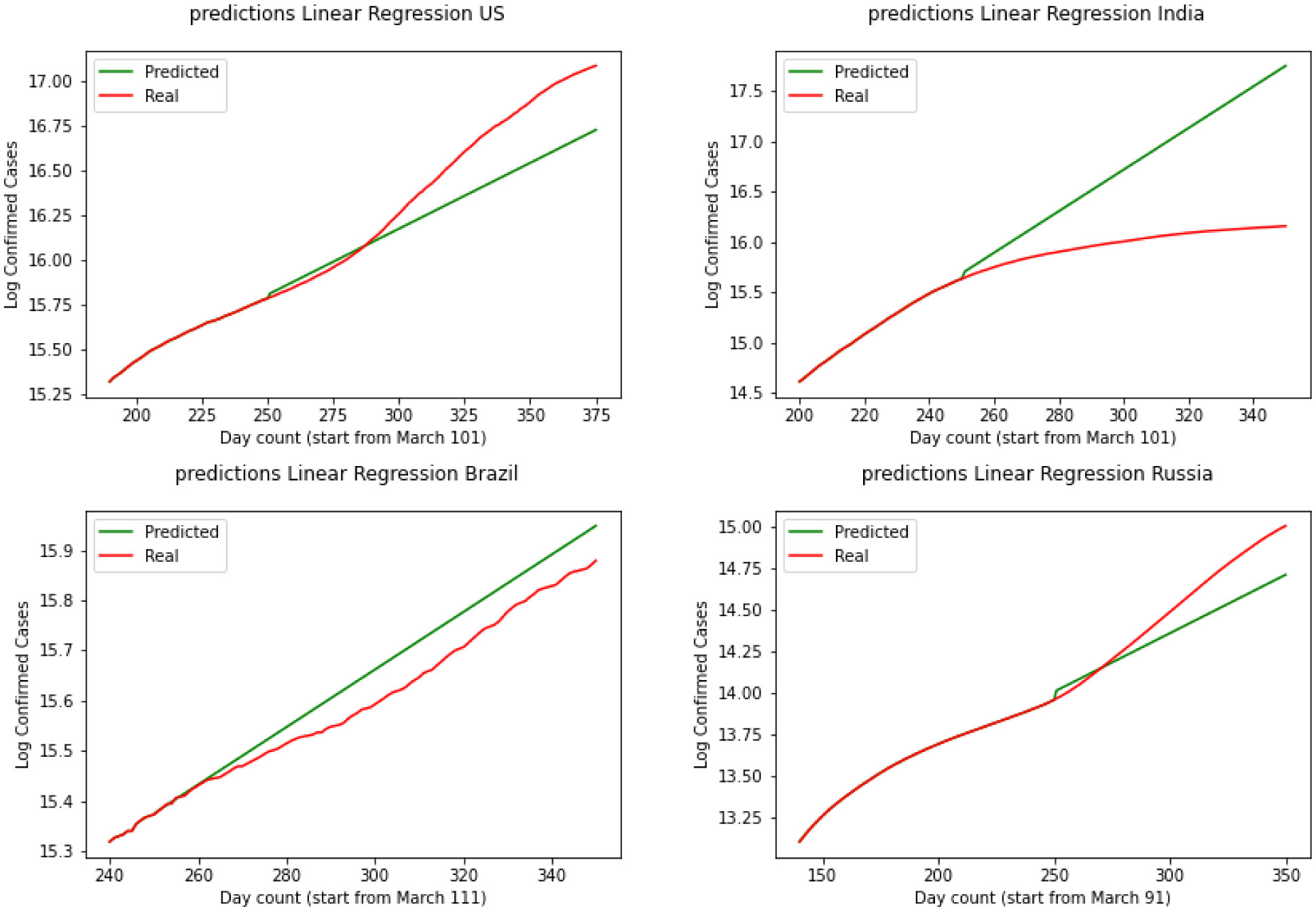
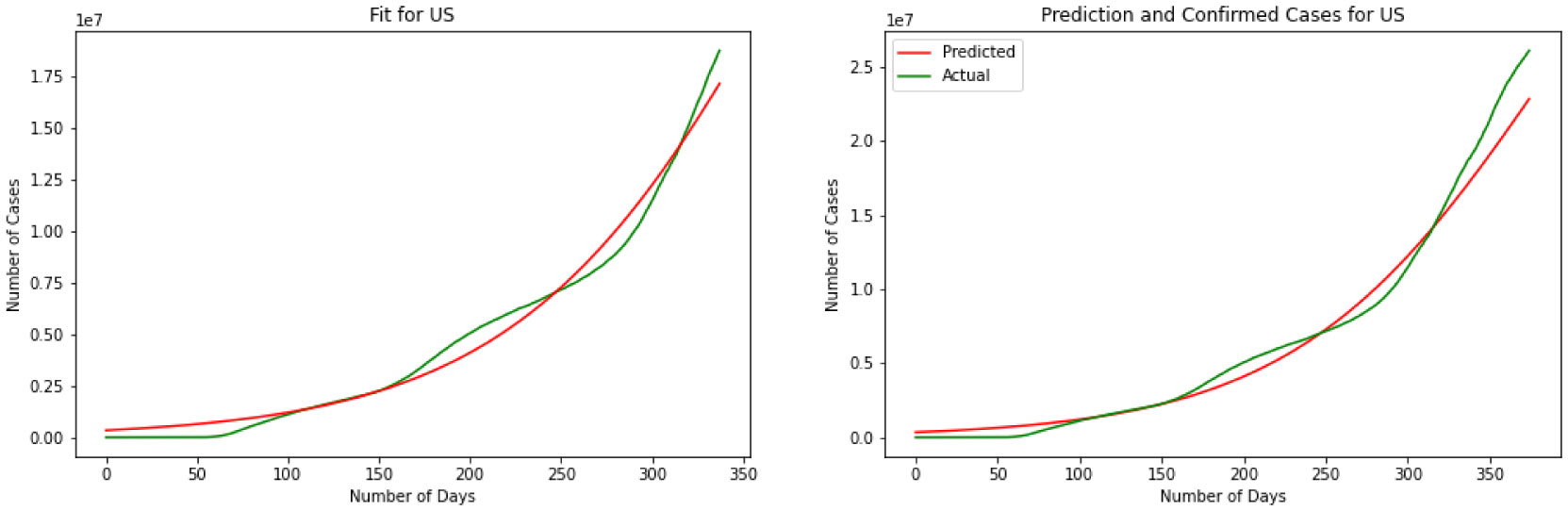
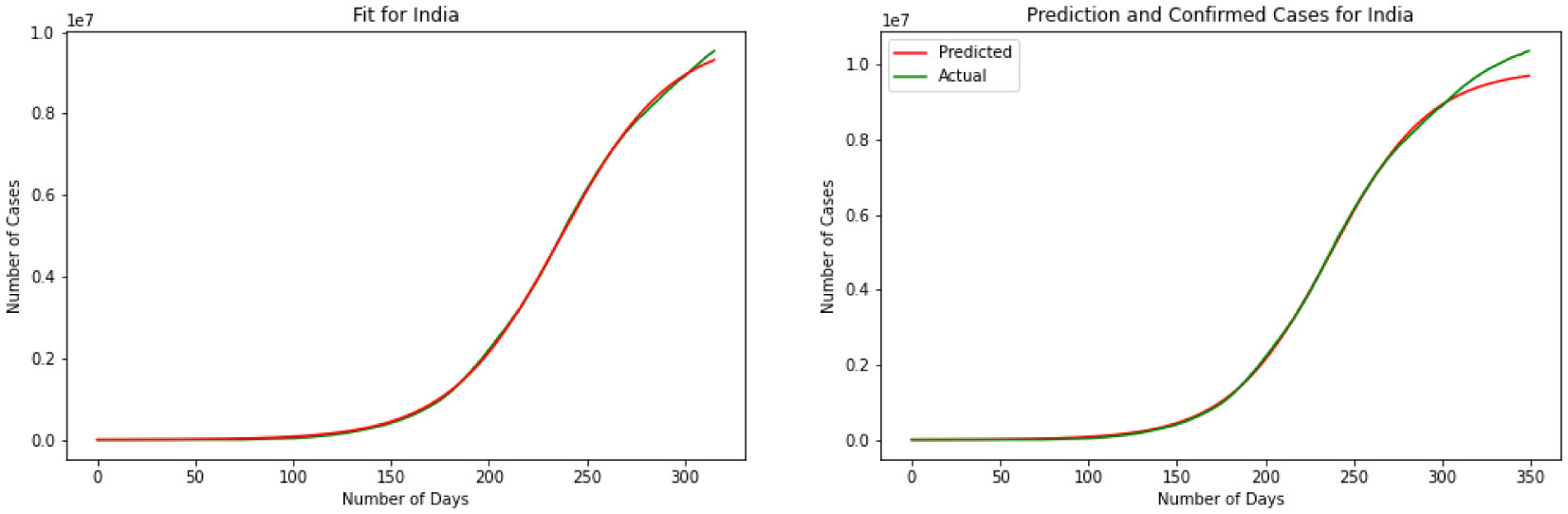
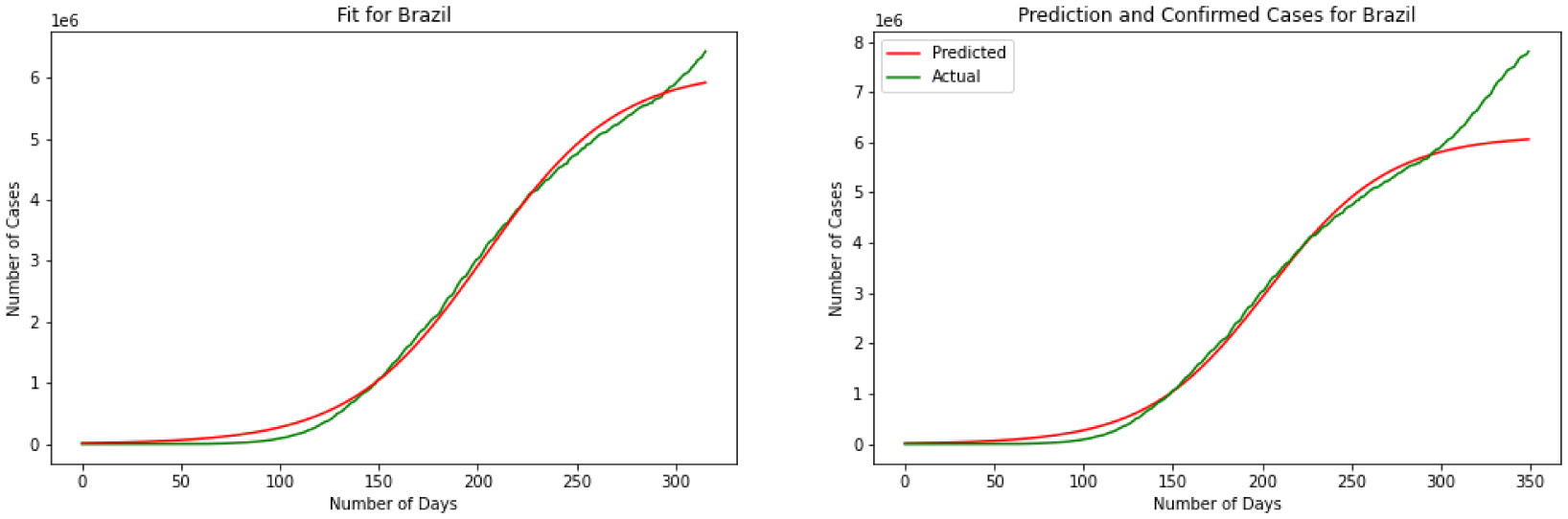
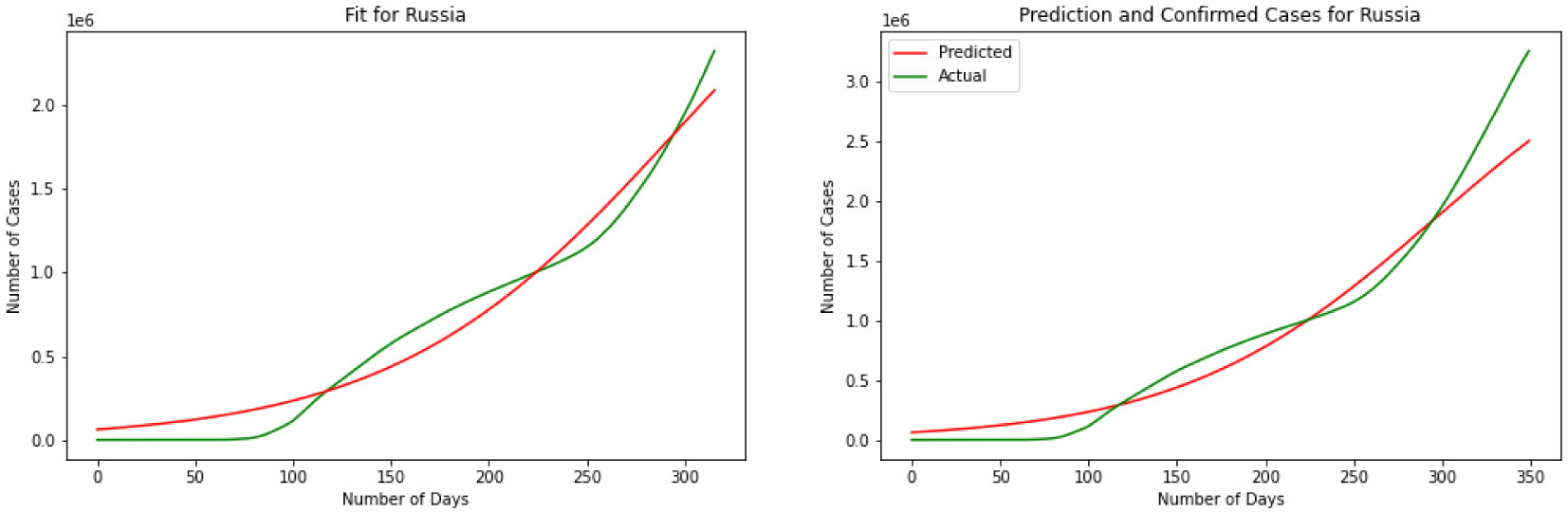
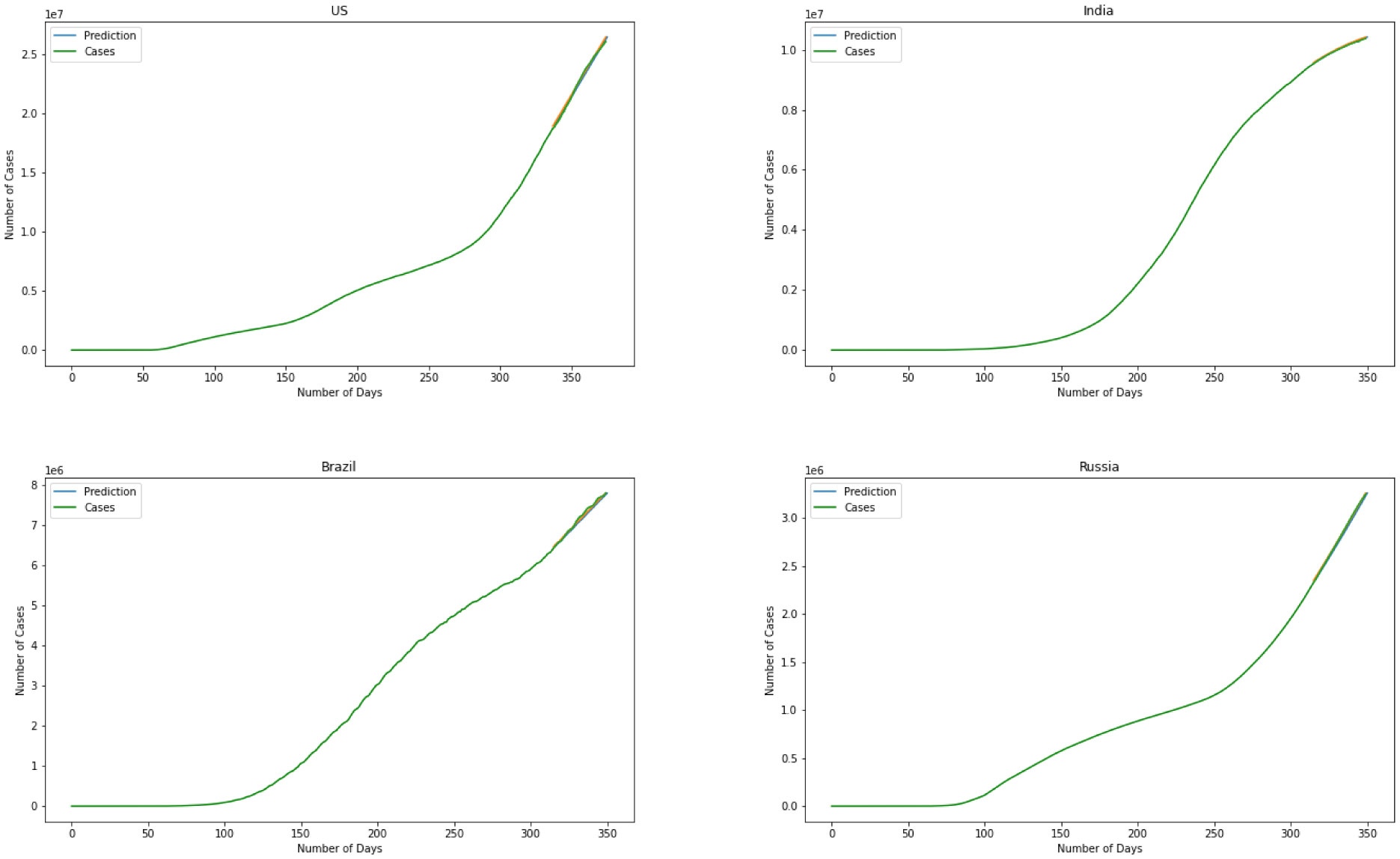
Figures(11)

Saina Abolmaali, Samira Shirzaei. A comparative study of SIR Model, Linear Regression, Logistic Function and ARIMA Model for forecasting COVID-19 cases[J]. AIMS Public Health, 2021, 8(4): 598-613. doi: 10.3934/publichealth.2021048







 DownLoad:
DownLoad: