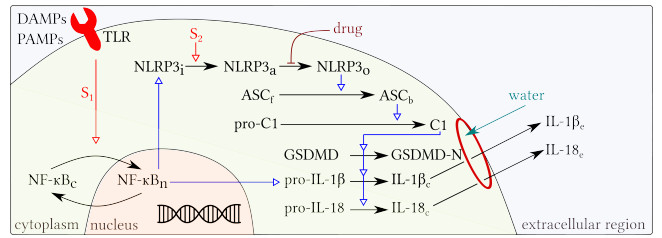
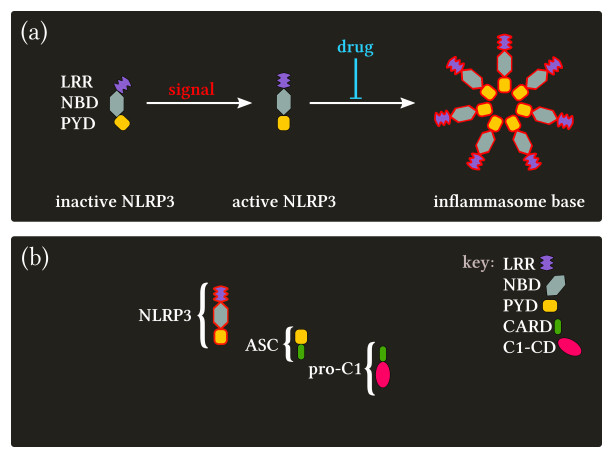
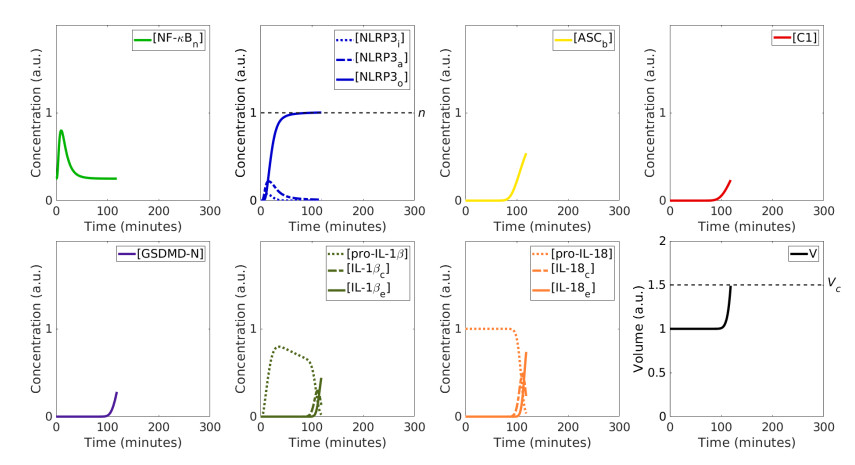
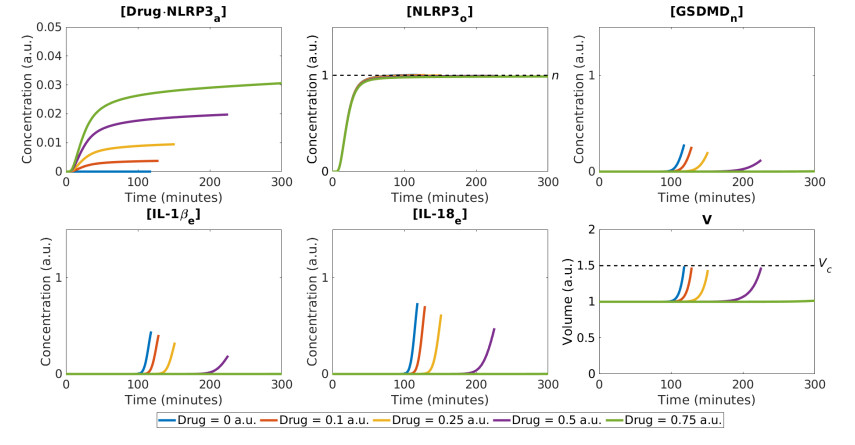
Pyroptosis is an inflammatory mode of cell death that can contribute to the cytokine storm associated with severe cases of coronavirus disease 2019 (COVID-19). The formation of the NLRP3 inflammasome is central to pyroptosis, which may be induced by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Inflammasome formation, and by extension pyroptosis, may be inhibited by certain anti-inflammatory drugs. In this study, we present a single-cell mathematical model that captures the formation of the NLRP3 inflammasome, pyroptotic cell death and responses to anti-inflammatory intervention that hinder the formation of the NLRP3 inflammasome. The model is formulated in terms of a system of ordinary differential equations (ODEs) that describe the dynamics of the key biological components involved in pyroptosis. Our results demonstrate that an anti-inflammatory drug can delay the formation of the NLRP3 inflammasome, and thus may alter the mode of cell death from inflammatory (pyroptosis) to non-inflammatory (e.g., apoptosis). The single-cell model is implemented within a SARS-CoV-2 tissue simulator, in collaboration with a multidisciplinary coalition investigating within host-dynamics of COVID-19. In this paper, we additionally provide an overview of the SARS-CoV-2 tissue simulator and highlight the effects of pyroptosis on a cellular level.
Citation: Sara J Hamis, Fiona R Macfarlane. A single-cell mathematical model of SARS-CoV-2 induced pyroptosis and the effects of anti-inflammatory intervention[J]. AIMS Mathematics, 2021, 6(6): 6050-6086. doi: 10.3934/math.2021356
Pyroptosis is an inflammatory mode of cell death that can contribute to the cytokine storm associated with severe cases of coronavirus disease 2019 (COVID-19). The formation of the NLRP3 inflammasome is central to pyroptosis, which may be induced by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Inflammasome formation, and by extension pyroptosis, may be inhibited by certain anti-inflammatory drugs. In this study, we present a single-cell mathematical model that captures the formation of the NLRP3 inflammasome, pyroptotic cell death and responses to anti-inflammatory intervention that hinder the formation of the NLRP3 inflammasome. The model is formulated in terms of a system of ordinary differential equations (ODEs) that describe the dynamics of the key biological components involved in pyroptosis. Our results demonstrate that an anti-inflammatory drug can delay the formation of the NLRP3 inflammasome, and thus may alter the mode of cell death from inflammatory (pyroptosis) to non-inflammatory (e.g., apoptosis). The single-cell model is implemented within a SARS-CoV-2 tissue simulator, in collaboration with a multidisciplinary coalition investigating within host-dynamics of COVID-19. In this paper, we additionally provide an overview of the SARS-CoV-2 tissue simulator and highlight the effects of pyroptosis on a cellular level.

| [1] |
P. Zhou, X. Yang, X. Wang, B. Hu, L. Zhang, W. Zhang, et al., A pneumonia outbreak associated with a new coronavirus of probable bat origin, Nature, 579 (2020), 270–273. doi: 10.1038/s41586-020-2012-7

|
| [2] |
J. K. Y. Yap, M. Moriyama, A. Iwasaki, Inflammasomes and pyroptosis as therapeutic targets for COVID-19, J. Immunol., 205 (2020), 307–312. doi: 10.4049/jimmunol.2000513
|
| [3] | M. Z. Tay, C. M. Poh, L. Rénia, P. A. MacAry, L. F. P. Ng, The trinity of COVID-19: Immunity, inflammation and intervention, Nat. Rev. Immunol., 579 (2020), 363–374. |
| [4] |
M. Soy, G. Keser, P. Atagündüz, F. Tabak, I. Atagündüz, S. Kayhan, Cytokine storm in COVID-19: Pathogenesis and overview of anti-inflammatory agents used in treatment, Clin. Rheumatol., 39 (2020), 2085–2094. doi: 10.1007/s10067-020-05190-5
|
| [5] |
D. Tang, P. Comish, R. Kang, The hallmarks of COVID-19 disease, PLoS Pathog., 16 (2020), e1008536. doi: 10.1371/journal.ppat.1008536
|
| [6] |
A. Shah, Novel coronavirus-induced NLRP3 inflammasome activation: A potential drug target in the treatment of COVID-19, Front. Immunol., 11 (2020), 1021. doi: 10.3389/fimmu.2020.01021
|
| [7] | M. Z. Ratajczak, M. Kucia, SARS-CoV-2 infection and overactivation of NLRP3 inflammasome as a trigger of cytokine storm and risk factor for damage of hematopoietic stem cells, Leukemia, 4 (2020), 1726–1729. |
| [8] |
Y. Fu, Y. Cheng, Y. Wu, Understanding SARS-CoV-2-mediated inflammatory responses: From mechanisms to potential therapeutic tools, Virol. Sin., 35 (2020), 266–271. doi: 10.1007/s12250-020-00207-4
|
| [9] |
S. Nagashima, M. C. Mendes, A. P. Camargo, N. H. Borges, T. M. Godoy, A. F. R. Miggiolaro, et al., Endothelial dysfunction and thrombosis in patients with COVID-19-brief report, Arterioscler. Thromb. Vasc. Biol., 40 (2020), 2404–2407. doi: 10.1161/ATVBAHA.120.314860
|
| [10] |
Y. Jamilloux, T. Henry, A. Belot, S. Viel, M. Fauter, T. El Jammal, et al., Should we stimulate or suppress immune responses in COVID-19: Cytokine and anti-cytokine interventions, Autoimmun. Rev., 19 (2020), 102567. doi: 10.1016/j.autrev.2020.102567
|
| [11] |
C. Y. Taabazuing, M. C. Okondo, D. A. Bachovchin, Pyroptosis and apoptosis pathways engage in bidirectional crosstalk in monocytes and macrophages, Cell Chem. Biol., 24 (2017), 507–514. doi: 10.1016/j.chembiol.2017.03.009
|
| [12] |
N. Kelley, D. Jeltema, Y. Duan, Y. He, The NLRP3 inflammasome: An overview of mechanisms of activation and regulation, Int. J. Mol. Sci., 20 (2019), 3328. doi: 10.3390/ijms20133328
|
| [13] |
T. Bergsbaken, S. L. Fink, B. T. Cookson, Pyroptosis: Host cell death and inflammation, Nat. Rev. Microbiol., 7 (2009), 99–109. doi: 10.1038/nrmicro2070
|
| [14] |
C. A. Dinarello, Immunological and inflammatory functions of the interleukin-1 family, Annu. Rev. Immunol., 27 (2009), 519–550. doi: 10.1146/annurev.immunol.021908.132612
|
| [15] |
Y. He, H. Hara, G. Núñez, Mechanism and regulation of NLRP3 inflammasome activation, Trends Biochem. Sci., 41 (2016), 1012–1021. doi: 10.1016/j.tibs.2016.09.002
|
| [16] |
Z. B. Zalinger, R. Elliott, S.R. Weiss, Role of the inflammasome-related cytokines IL-1 and IL-18 during infection with murine coronavirus, J. Neurovirol., 23 (2017), 845–854. doi: 10.1007/s13365-017-0574-4
|
| [17] |
A. Stutz, D. T. Golenbock, E. Latz, Inflammasomes: Too big to miss, J. Clin. Invest., 119 (2009), 3502–3511. doi: 10.1172/JCI40599
|
| [18] | J. J. O'Shea, M. Gadina, R. M. Siegel, J. Farber, Cytokines, in Rheumatology, (2015), 99–112. |
| [19] |
P. Song, W. Li, J. Xie, Y. Hou, C. You, Cytokine storm induced by SARS-CoV-2, Clin. Chim. Acta, 509 (2020), 280–287. doi: 10.1016/j.cca.2020.06.017
|
| [20] |
C. Huang, Y. Wang, X. Li, L. Ren, J. Zhao, Y. Hu, et al., Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China, The Lancet, 395 (2020), 497–506. doi: 10.1016/S0140-6736(20)30183-5
|
| [21] |
M. Bertinaria, S. Gastaldi, E. Marini, M. Giorgis, Development of covalent NLRP3 inflammasome inhibitors: Chemistry and biological activity, Arch. Biochem. Biophys., 670 (2019), 116–139. doi: 10.1016/j.abb.2018.11.013
|
| [22] |
K. Tsuchiya, Inflammasome-associated cell death: Pyroptosis, apoptosis, and physiological implications, Microbiol. Immunol., 64 (2020), 252–269. doi: 10.1111/1348-0421.12771
|
| [23] |
S. L. Fink, B. T. Cookson, Apoptosis, pyroptosis, and necrosis: Mechanistic description of dead and dying eukaryotic cells, Infect. Immun., 73 (2005), 1907–1916. doi: 10.1128/IAI.73.4.1907-1916.2005
|
| [24] | A. G. Kozloski, Inflammasome, Mater. Methods, 10 (2020), 2869. |
| [25] |
S. Christgen, D. E. Place, T. D. Kanneganti, Toward targeting inflammasomes: Insights into their regulation and activation, Cell Res., 30 (2020), 315–327. doi: 10.1038/s41422-020-0295-8
|
| [26] |
Z. Zheng, G. Li, Mechanisms and therapeutic regulation of pyroptosis in inflammatory diseases and cancer, Int. J. Mol. Sci., 21 (2020), 1456. doi: 10.3390/ijms21041456
|
| [27] |
M. G. Sanders, M. J. Parsons, A. G. Howard, J. Liu, S. R. Fassio, J. A. Martinez, et al., Single-cell imaging of inflammatory caspase dimerization reveals differential recruitment to inflammasomes, Cell Death Dis., 6 (2015), e1813. doi: 10.1038/cddis.2015.186
|
| [28] |
C. Semino, S. Carta, M. Gattorno, R. Sitia, A. Rubartelli, Progressive waves of IL-1$\beta$ release by primary human monocytes via sequential activation of vesicular and gasdermin D-mediated secretory pathways, Cell Death Dis., 9 (2018), 1–14. doi: 10.1038/s41419-017-0012-9
|
| [29] |
G. Lopez-Castejon, D. Brough, Understanding the mechanism of IL-1$\beta$ secretion, Cytokine Growth Factor Rev., 22 (2011), 189–195. doi: 10.1016/j.cytogfr.2011.10.001
|
| [30] | P. Broz, P. Pelegrín, F. Shao, The gasdermins, a protein family executing cell death and inflammation, Nat. Rev. Immunol., 20 (2019), 143–157. |
| [31] | L. DiPeso, D. X. Ji, R. E. Vance, J. V. Price, Cell death and cell lysis are separable events during pyroptosis, Cell Death Dis., 3 (2017), 1–10. |
| [32] |
D. Brough, N. J. Rothwell, Caspase-1-dependent processing of pro-interleukin-1$\beta$ is cytosolic and precedes cell death, J. Cell Sci., 120 (2007), 772–781. doi: 10.1242/jcs.03377
|
| [33] |
K. Schleich, I. N. Lavrik, Mathematical modeling of apoptosis, Cell Comm. Signal., 11 (2013), 1–7. doi: 10.1186/1478-811X-11-1
|
| [34] |
S. L. Spencer, P. K. Sorger, Measuring and modeling apoptosis in single cells, Cell, 144 (2011), 926–939. doi: 10.1016/j.cell.2011.03.002
|
| [35] |
W. Wang, T. Zhang, Caspase-1-mediated pyroptosis of the predominance for driving CD4 T cells death: A nonlocal spatial mathematical model, Bull. Math. Biol., 80 (2018), 540–582. doi: 10.1007/s11538-017-0389-8
|
| [36] |
D. Veltman, T. Laeremans, E. Passante, H. J. Huber, Signal transduction analysis of the NLRP3-inflammasome pathway after cellular damage and its paracrine regulation, J. Theor. Biol., 415 (2017), 125–136. doi: 10.1016/j.jtbi.2016.12.016
|
| [37] | Y. Bozkurt, A. Demir, B. Erman, A. Gül, Unified modeling of familial mediterranean fever and cryopyrin associated periodic syndromes, Comp. Math. Meth. Med., 15 (2015), 893507. |
| [38] | F. López-Caamal, H. J. Huber, Stable IL-$1\beta$-activation in an inflammasome signalling model depends on positive and negative-feedbacks and tight regulation of protein production, IEEE ACM T. Comput. Bi., 16 (2018), 627–637. |
| [39] | WHO, World Health Organisation Statement on the Pandemic, 2020. Available from: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19–-11-march-2020. |
| [40] |
F. Akpinar, B. Inankur, J. Yin, Spatial-temporal patterns of viral amplification and interference initiated by a single infected cell, J. Virol., 90 (2016), 7552–7566. doi: 10.1128/JVI.00807-16
|
| [41] |
A. Bankhead, E. Mancini, A. C. Sims, R. S. Baric, S. McWeeney, P. M. A. Sloot, A simulation framework to investigate in vitro viral infection dynamics, J. Comput. Sci., 4 (2013), 127–134. doi: 10.1016/j.jocs.2011.08.007
|
| [42] |
A. L. Bauer, C. Beauchemin, A. S. Perelson, Agent-based modeling of host–pathogen systems: The successes and challenges, Inf. Sci., 179 (2009), 1379–1389. doi: 10.1016/j.ins.2008.11.012
|
| [43] |
G. Bocharov, A. Meyerhans, N. Bessonov, S. Trofimchuk, V. Volpert, Spatiotemporal dynamics of virus infection spreading in tissues, PLoS One, 11 (2016), e0168576. doi: 10.1371/journal.pone.0168576
|
| [44] | C. Beauchemin, S. Forrest, F. T. Koster, Modeling influenza viral dynamics in tissue, in International Conference on Artificial Immune Systems (Springer), (2006), 23–36. |
| [45] |
C. Beauchemin, Probing the effects of the well-mixed assumption on viral infection dynamics, J. Theor. Biol., 242 (2006), 464–477. doi: 10.1016/j.jtbi.2006.03.014
|
| [46] |
C. Beauchemin, J. Samuel, J. Tuszynski, A simple cellular automaton model for influenza A viral infections, J. Theor. Biol., 232 (2005), 223–234. doi: 10.1016/j.jtbi.2004.08.001
|
| [47] |
D. Levin, S. Forrest, S. Banerjee, C. Clay, J. Cannon, M. Moses, et al., A spatial model of the efficiency of T cell search in the influenza-infected lung, J. Theor. Biol., 398 (2016), 52–63. doi: 10.1016/j.jtbi.2016.02.022
|
| [48] | N. Fachada, V. V. Lopes, A. Rosa, Simulating antigenic drift and shift in influenza A, in Proceedings of the 2009 ACM symposium on Applied Computing, (2009), 2093–2100. |
| [49] |
A. L. Jenner, F. Frascoli, A. C. F. Coster, P. S. Kim, Enhancing oncolytic virotherapy: Observations from a Voronoi cell-based model, J. Theor. Biol., 485 (2020), 110052. doi: 10.1016/j.jtbi.2019.110052
|
| [50] |
D. Wodarz, A. Hofacre, J. W. Lau, Z. Sun, H. Fan, N. L. Komarova, Complex spatial dynamics of oncolytic viruses in vitro: Mathematical and experimental approaches, PLoS Comput. Biol., 8 (2012), e1002547. doi: 10.1371/journal.pcbi.1002547
|
| [51] |
G. An, Introduction of an agent-based multi-scale modular architecture for dynamic knowledge representation of acute inflammation, Theor. Biol. Med., 5 (2008), 11. doi: 10.1186/1742-4682-5-11
|
| [52] |
R. C. Cockrell, G. An, Examining the controllability of sepsis using genetic algorithms on an agent-based model of systemic inflammation, PLoS Comput. Biol., 14 (2018), e1005876. doi: 10.1371/journal.pcbi.1005876
|
| [53] | F. Fatehi, R. J. Bingham, E. C. Dykeman, P. G. Stockley, R. Twarock, Comparing antiviral strategies against COVID-19 via multi-scale within host modelling, preprint, arXiv: 2010.08957. |
| [54] |
T. J. Sego, J. O. Aponte-Serrano, J. F. Gianlupi, S. R. Heaps, K. Breithaupt, L. Brusch, et al., A modular framework for multiscale, multicellular, spatiotemporal modeling of acute primary viral infection and immune response in epithelial tissues and its application to drug therapy timing and effectiveness, PLoS Comput. Biol., 16 (2020), e1008451. doi: 10.1371/journal.pcbi.1008451
|
| [55] | Y. Wang, G. An, A. Becker, C. Cockrell, N. Collier, M. Craig, et al., Rapid community-driven development of a SARS-CoV-2 tissue simulator, preprint, bioR$\chi$iv: 2020.04.02.019075. Available from: https://www.biorxiv.org/content/10.1101/2020.04.02.019075v3. |
| [56] | A. V. Bagaev, A. Y. Garaeva, E. S. Lebedeva, A. V. Pichugin, R. I. Ataullakhanov, F. I. Ataullakhanov, Elevated pre-activation basal level of nuclear NF-$\kappa$B in native macrophages accelerates LPS-induced translocation of cytosolic NF-$\kappa$B into the cell nucleus, Sci. Reports, 9 (2019), 1–16. |
| [57] |
C. Zhang, C. Zhao, X. Chen, R. Tao, S. Wang, G. Meng, et al., Induction of ASC pyroptosis requires gasdermin D or caspase-1/11-dependent mediators and IFN$\beta$ from pyroptotic macrophages, Cell Death Dis., 11 (2020), 470. doi: 10.1038/s41419-020-2664-0
|
| [58] | Y. Huang, H. Jiang, Y. Chen, X. Wang, Y. Yang, J. Tao, et al., Tranilast directly targets NLRP3 to treat inflammasome-driven diseases, EMBO Mol. Med., 10 (2018), e8689. |
| [59] |
M. S. Salahudeen, P. S. Nishtala, An overview of pharmacodynamic modelling, ligand-binding approach and its application in clinical practice, Saudi Pharm. J., 25 (2017), 165–175. doi: 10.1016/j.jsps.2016.07.002
|
| [60] |
N. M. de Vasconcelos, N. Van Opdenbosch, H. Van Gorp, E. Parthoens, M. Lamkanfi, Single-cell analysis of pyroptosis dynamics reveals conserved GSDMD-mediated subcellular events that precede plasma membrane rupture, Cell Death Diff., 26 (2019), 146–161. doi: 10.1038/s41418-018-0106-7
|
| [61] |
S. Han, T. B. Lear, J. A. Jerome, S. Rajbhandari, C. A. Snavely, D. L. Gulick, et al., Lipopolysaccharide primes the NALP3 inflammasome by inhibiting its ubiquitination and degradation mediated by the SCFFBXL2 E3 ligase, J. Bio. Chem., 290 (2015), 18124–18133. doi: 10.1074/jbc.M115.645549
|
| [62] | J. Cheng, A. L. Waite, E. R., Tkaczyk, K. Ke, N. Richards, A. J. Hunt, et al., Kinetic properties of ASC protein aggregation in epithelial cells, J. Cell. Physiol., 222 (2010), 738–747. |
| [63] |
J. Ruland, Inflammasome: Putting the pieces together, Cell, 156 (2014), 1127–1129. doi: 10.1016/j.cell.2014.02.038
|
| [64] |
J. Chai, Y. Shi, Apoptosome and inflammasome: Conserved machineries for caspase activation, Nat. Sci. Rev., 1 (2014), 101–118. doi: 10.1093/nsr/nwt025
|
| [65] |
A. Iliev, N. Kyurkchiev, S. Markov, On the approximation of the step function by some sigmoid functions, Math. Comput. Simul., 133 (2017), 223–234. doi: 10.1016/j.matcom.2015.11.005
|
| [66] | MATLAB, version 1.8.0_202 (R2019n). The MathWorks Inc., Natick, Massachusetts, 2019. |
| [67] | M. A. Moors, S. B. Mizel, Proteasome-mediated regulation of interleukin-1$\beta$ turnover and export in human monocytes, J. Leukocyte Biol., 68 (2000), 131–136. |
| [68] |
F. Martín-Sánchez, C. Diamond, M. Zeitler, A. I. Gomez, A. Baroja-Mazo, J. Bagnall, et al., Inflammasome-dependent IL-1$\beta$ release depends upon membrane permeabilisation, Cell Death Diff., 23 (2016), 1219–1231. doi: 10.1038/cdd.2015.176
|
| [69] |
G. Qian, A. Mahdi, Sensitivity analysis methods in the biomedical sciences, Math Biosci., 323 (2020), 108306. doi: 10.1016/j.mbs.2020.108306
|
| [70] | S. Hamis, S. Stratiev, G. G. Powathil, Uncertainty and sensitivity analyses methods for agent-based mathematical models: An introductory review, in The Physics of Cancer: Research Advances (ed. Bernard Gerstman), Singapore: World Scientific Publishing, 2021. |
| [71] |
R. P. Dickinson, R. J. Gelinas, Sensitivity analysis of ordinary differential equation systems - A direct method, J. Comput. Phys., 21 (1976), 123–143. doi: 10.1016/0021-9991(76)90007-3
|
| [72] |
A. Ghaffarizadeh, R. Heiland, S. H. Friedman, S. M. Mumenthaler, P. Macklin, PhysiCell: An open source physics-based cell simulator for 3-D multicellular systems, PLoS Comput. Biol., 14 (2018), e1005991. doi: 10.1371/journal.pcbi.1005991
|
| [73] |
K. Lipinska, K. E. Malone, M. Moerland, C. Kluft, Applying caspase-1 inhibitors for inflammasome assays in human whole blood, J. Immunol. Meth., 411 (2014), 66–69. doi: 10.1016/j.jim.2014.05.018
|
| [74] | K. Schroder, J. Tschopp, The inflammasomes, Cell, 140 (2010), 821–832. |
| [75] |
W. He, H. Wan, L. Hu, P. Chen, X. Wang, Z. Huang, et al., Gasdermin D is an executor of pyroptosis and required for interleukin-1$\beta$ secretion, Cell Res., 25 (2015), 1285–1298. doi: 10.1038/cr.2015.139
|
| [76] | S. Lee, M. Hirohama, M. Noguchi, K. Nagata, A. Kawaguchi, Influenza A virus infection triggers pyroptosis and apoptosis of respiratory epithelial cells through the type Ⅰ interferon signaling pathway in a mutually exclusive manner, J. Virol., 92 (2018), e00396-18. |
| [77] |
M. P. Lythgoe, P. Middleton, Ongoing clinical trials for the management of the COVID-19 pandemic, Trends Pharmacol. Sci., 41 (2020), 363–382. doi: 10.1016/j.tips.2020.03.006
|
| [78] |
K. Sharun, R. Tiwari, J. Dhama, K. Dhama, Dexamethasone to combat cytokine storm in COVID-19: Clinical trials and preliminary evidence, Int. J. Surg., 82 (2020), 179–181. doi: 10.1016/j.ijsu.2020.08.038
|
| [79] |
RECOVERY Collaborative Group, Dexamethasone in hospitalized patients with covid-19: Preliminary report, N. Engl. J. Med., 384 (2020), 693–704. doi: 10.1056/NEJMoa2021436
|
| [80] |
T. Rhen, J. A. Cidlowski, Antiinflammatory action of glucocorticoids: New mechanisms for old drugs, N. Engl. J. Med., 353 (2005), 1711–1723. doi: 10.1056/NEJMra050541
|
| [81] |
M. Cornut, E. Bourdonnay, H. Thomas, Transcriptional Regulation of Inflammasomes, Int. J. Mol. Sci., 21 (2020), 8087. doi: 10.3390/ijms21218087
|
| [82] |
S. Christgen, D. E. Place, T. D. Kanneganti, Toward targeting inflammasomes: Insights into their regulation and activation, Cell Res., 30 (2020), 315–327. doi: 10.1038/s41422-020-0295-8
|
| [83] |
F. Chen, G. Jiang, H. Liu, Z. Li, Y. Pei, H. Wang, et al., Melatonin alleviates intervertebral disc degeneration by disrupting the IL-1$\beta$/NF-$\kappa$B-NLRP3 inflammasome positive feedback loop, Bone Res., 8 (2020), 1–13. doi: 10.1038/s41413-019-0078-3
|
| [84] |
H. Wu, C. Zhao, Q. Xie, J. Xu, G. Fei, TLR2-melatonin feedback loop regulates the activation of NLRP3 inflammasome in murine allergic airway inflammation, Front. Immunol., 11 (2020), 172. doi: 10.3389/fimmu.2020.00172
|
| [85] |
B. B. Mishra, V. A. K. Rathinam, G. W. Martens, A. J. Martinot, H. Kornfeld, K. A. Fitzgerald, et al., Nitric oxide controls the immunopathology of tuberculosis by inhibiting NLRP3 inflammasome–dependent processing of IL-1$\beta$, Nat. Immunol., 14 (2013), 52–60. doi: 10.1038/ni.2474
|
| [86] |
G. Guarda, M. Braun, F. Staehli, A. Tardivel, C. Mattmann, I. Förster, ., Type Ⅰ interferon inhibits interleukin-1 production and inflammasome activation, Immunity, 34 (2011), 213–223. doi: 10.1016/j.immuni.2011.02.006
|
| [87] | K. G. Lokugamage, A. Hage, M. de Vries, A. M. Valero-JImenez, C. Schindewolf, M. Dittmann, et al., Type Ⅰ interferon susceptibility distinguishes SARS-CoV-2 from SARS-CoV, J. Virol., 94 (2020), e01410-20. |
| [88] |
C. Juliana, T. Fernandes-Alnemri, S. Kang, A. Farias, F. Qin, E. S. Alnemri, Non-transcriptional priming and deubiquitination regulate NLRP3 inflammasome activation, J. Biol. Chem., 287 (2012), 36617–36622. doi: 10.1074/jbc.M112.407130
|
| [89] |
J. S. Lolkema, D. Slotboom, The Hill analysis and co-ion–driven transporter kinetics, J. Gen. Physio., 145 (2015), 565–574. doi: 10.1085/jgp.201411332
|
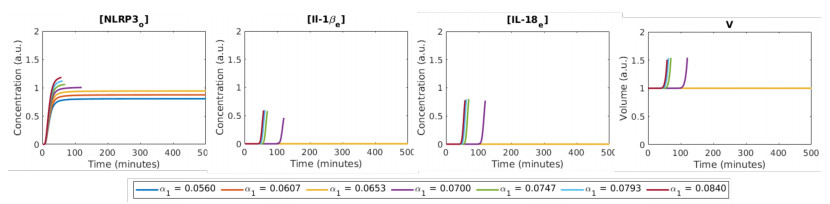
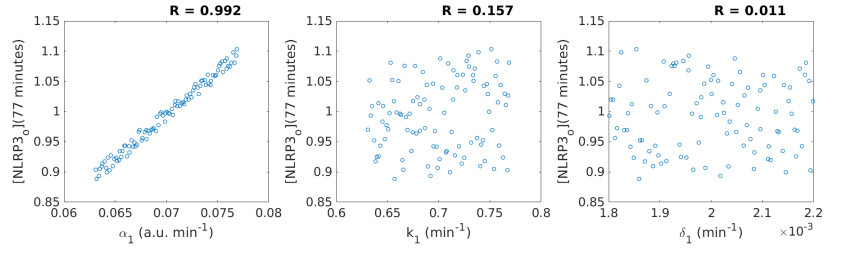
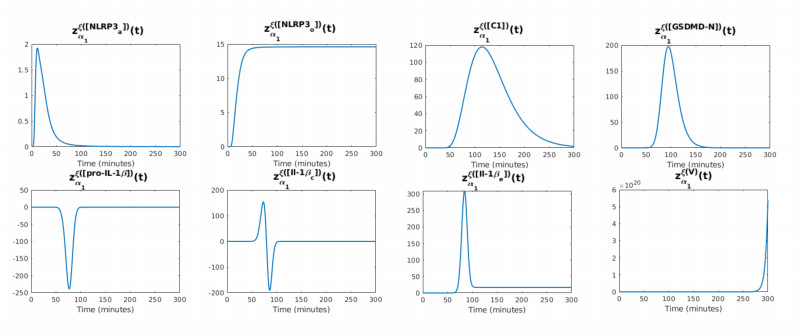
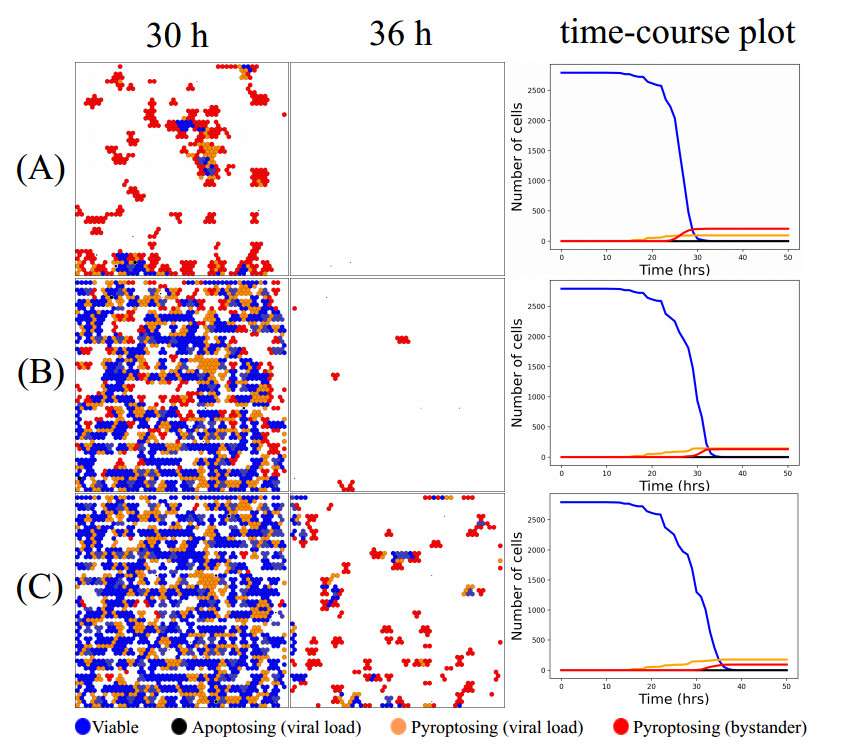
Figures(12) / Tables(2)

Sara J Hamis, Fiona R Macfarlane. A single-cell mathematical model of SARS-CoV-2 induced pyroptosis and the effects of anti-inflammatory intervention[J]. AIMS Mathematics, 2021, 6(6): 6050-6086. doi: 10.3934/math.2021356







 DownLoad:
DownLoad: