Citation: Solomon A. A., Michel Child, Upeksha Caldera, Christian Breyer. Exploiting wind-solar resource complementarity to reduce energy storage need[J]. AIMS Energy, 2020, 8(5): 749-770. doi: 10.3934/energy.2020.5.749

| [1] |
Solomon AA (2019) Large scale photovoltaics and the future energy system requirement. AIMS Energy 7: 600-618. doi: 10.3934/energy.2019.5.600

|
| [2] |
Denholm P, Margolis RM (2007) Evaluating the limits of solar photovoltaics (PV) in electric power systems utilizing energy storage and other enabling technologies. Energy Policy 35: 4424-4433. doi: 10.1016/j.enpol.2007.03.004
|
| [3] |
Lund PD, Lindgren J, Mikkola J, et al. (2015) Review of energy system flexibility measures to enable high levels of variable renewable electricity. Renewable Sustainable Energy Rev 45: 785-807. doi: 10.1016/j.rser.2015.01.057
|
| [4] |
Konziella H, Bruckner T (2016) Flexibility requirements of renewable energy based electricity systems-a review of research results and methodologies. Renewable Sustainable Energy Rev 53: 10-22. doi: 10.1016/j.rser.2015.07.199
|
| [5] |
Heide DM, Greiner Bremen L, Hoffmann C (2011) Reduced storage and balancing needs in a fully renewable European power system with excess wind and solar power generation. Renewable Energy 36: 2515-2523. doi: 10.1016/j.renene.2011.02.009
|
| [6] |
Solomon AA, Kammen DM, Callaway D (2016) Investigating the impact of wind-solar complementarities on energy storage requirement and the corresponding supply reliability criteria. Appl Energ 168: 130-145. doi: 10.1016/j.apenergy.2016.01.070
|
| [7] |
Heide D, Bremen L, Greiner M, et al. (2010) Seasonal optimal mix of wind and solar power in a future, highly renewable Europe. Renewable Energy 35: 2483-2489. doi: 10.1016/j.renene.2010.03.012
|
| [8] |
Esteban M, Zhang Q, Utama A (2012) Estimation of the energy storage requirement of a future 100% renewable energy system in Japan. Energy Policy 47: 22-31. doi: 10.1016/j.enpol.2012.03.078
|
| [9] | Budischak C, Sewell D, Thomson H, et al. (2013) Cost-minimized combinations of wind power, solar power and electrochemical storage, powering the grid up to 99.9% of the time. J Power Sources 225: 60-74. |
| [10] |
Child M, Breyer C (2016) The role of energy storage solutions in a 100% renewable Finnish energy system. Energy Procedia 99: 25-34. doi: 10.1016/j.egypro.2016.10.094
|
| [11] | Caldera U, Bogdanov D, Afanasyeva S, et al. (2016) Integration of reverse osmosis seawater desalination in the power sector, based on PV and wind energy, for the Kingdom of Saudi Arabia. Proceedings of 32nd European Photovoltaic Solar Energy Conference, Munchen, 21-24. |
| [12] |
Bogdanov D, Breyer C (2016) North-East Asian Super Grid for 100% renewable energy supply: Optimal mix of energy technologies for electricity, gas and heat supply options. Energ Convers Manage 112: 176-190. doi: 10.1016/j.enconman.2016.01.019
|
| [13] |
Child M, Breyer C (2016) Vision and initial feasibility analysis of a recarbonised Finnish energy system. Renewable Sustainable Energy Rev 66: 517-536. doi: 10.1016/j.rser.2016.07.001
|
| [14] |
Babrowski S, Jochem P, Fichtner W (2016) Electricity storage systems in the future German energy sector-An optimization of the German electricity generation system until 2040 considering grid restrictions. Comput Oper Res 66: 228-240. doi: 10.1016/j.cor.2015.01.014
|
| [15] |
Mileva A, Nelson JH, Johnston J, et al. (2013) Sunshot solar power reduces costs and uncertainty in future low-carbon electricity systems. Environ Sci Technol 47: 9053-9060. doi: 10.1021/es401898f
|
| [16] |
Brouwer A, van den Broek M, Zappa W, et al. (2016) Least-cost options for integrating intermittent renewables in low-carbon power systems. Appl Energ 161: 48-74. doi: 10.1016/j.apenergy.2015.09.090
|
| [17] |
Solomon AA, Faiman D, Meron G (2010) Properties and uses of storage for enhancing the grid penetration of very large-scale photovoltaic systems. Energy Policy 38: 5208-5222. doi: 10.1016/j.enpol.2010.05.006
|
| [18] | Solomon AA, Faiman D, Meron G (2011) Appropriate storage for high-penetration grid-connected photovoltaic plants. Energy Policy 40: 335-344. |
| [19] |
Solomon AA, Kammen DM, Callaway D (2014) The role of large-scale energy storage design and dispatch in the power grid: a study of very high grid penetration of variable renewable resources. Appl Energ 134: 75-89. doi: 10.1016/j.apenergy.2014.07.095
|
| [20] |
Solomon AA, Faiman D, Meron G (2010) Grid matching of large-scale wind energy conversion systems, alone and in tandem with large-scale photovoltaic systems: An Israeli case study. Energy Policy 38: 7070-7081. doi: 10.1016/j.enpol.2010.07.026
|
| [21] |
Solomon AA, Faiman D, Meron G (2010) The effects on grid matching and ramping requirements, of single and distributed PV systems employing various fixed and sun-tracking technologies. Energy Policy 38: 5469-5481. doi: 10.1016/j.enpol.2010.02.056
|
| [22] |
Solomon AA, Faiman D, Meron G (2010) An energy-based evaluation of the matching possibilities of very large photovoltaic plants to the electricity grid: Israel as a case study. Energy Policy 38: 5457-5468. doi: 10.1016/j.enpol.2009.12.024
|
| [23] |
Kroposki B (2017) Integrating high level of variable renewable energy into electricity power systems. J Mod Power Syst Cle 5: 831-827. doi: 10.1007/s40565-017-0339-3
|
| [24] |
Solomon AA, Bogdanov D, Breyer C (2019) Curtailment-storage-penetration nexus in energy transition. Appl Energ 235: 1351-1368. doi: 10.1016/j.apenergy.2018.11.069
|
| [25] |
Miglietta MM, Huld T, Monforti-Ferrario F (2017) Local complementarity of wind and solar energy resources over Europe: An assessment study from a meteorological perspective. J Appl Meteorol Climatol 56: 217-234. doi: 10.1175/JAMC-D-16-0031.1
|
| [26] | Monferti F, Huld T, Bódis K, et al. (2014) Assessing complementarity of wind and solar resources for energy production in Italy. A Monte Carlo approach. Renewable Energy 63: 576-586. |
| [27] |
Hoicha, EC, Rowlands IH (2011) Solar and wind resource complementarity: Advancing options for renewable electricity integration in Ontario, Canada. Renewable Energy 36: 97-107. doi: 10.1016/j.renene.2010.06.004
|
| [28] | Gerlach AK, Stetter D, Schmid J, et al. (2011) PV and wind power-complementary technologies. 26th European Photovoltaic Solar Energy Conference, Hamburg, 5-9. |
| [29] |
Huber M, Dimkova D, Hamacher T (2014) Integration of wind and solar power in Europe: Assessment of flexibility requirements. Energy 69: 236-246. doi: 10.1016/j.energy.2014.02.109
|
| [30] |
Rosa CDOCS, Costa KA, Christo EDS, et al. (2017) Complementarity of hydro, photovoltaic, and wind power in Rio de Janeiro State. Sustainability 9: 1130. doi: 10.3390/su9071130
|
| [31] |
Lund H (2006) Large-scale integration of optimal combinations of PV, wind and wave power into the electricity supply. Renewable Energy 31: 503-515. doi: 10.1016/j.renene.2005.04.008
|
| [32] |
Slusarewicz JH, Cohan DS (2018) Assessing solar and wind complementarity in Texas. Renew Wind Water Solar 5: 7. doi: 10.1186/s40807-018-0054-3
|
| [33] | da Luz T, Moura P (2019) Power generation expansion planning with complementarity between renewable sources and regions for 100% renewable energy systems. Int T Electr Energy 29: e2817. |
| [34] |
Shaner MR, Davis SJ, Lewis NS, et al. (2018) Geophysical constraints on the reliability of solar and wind power in the United States. Energy Environ Sci 11: 914-925. doi: 10.1039/C7EE03029K
|
| [35] | Huang B, Krishnan V, Hodge BM (2018) Analyzing the impacts of variable renewable resources on California net-load ramp events. 2018 IEEE Power & Energy Society General Meeting, Portland, 5-10. |
| [36] |
Risso A, Beluco A, Alves RCM (2018) Complementarity roses evaluating spatial complementarity in time between energy resources. Energies 11: 1918. doi: 10.3390/en11071918
|
| [37] |
Min CG, Kim MK (2017) Impact of the complementarity between variable generation resources and load on the flexibility of the Korean power system. Energies 10: 1719. doi: 10.3390/en10111719
|
| [38] |
Santos-Alamillos FJ, Pozo-Vazques D, Ruiz-Arias JA, et al. (2015) Combining wind farms with concentrating solar plants to provide stable renewable power. Renewable Energy 76: 539-550. doi: 10.1016/j.renene.2014.11.055
|
| [39] |
Gallardo RP, Rios AM, Ramirez JS (2020) Analysis of the solar and wind energetic complementarity in Mexico. J Clean Prod 268: 122323. doi: 10.1016/j.jclepro.2020.122323
|
| [40] |
Han S, Zhang L, Liu Y, et al. (2019) Quantitative evaluation method for the complementarity of wind-solar-hydro power and optimization of wind-solar ratio. Appl Energ 236: 973-984. doi: 10.1016/j.apenergy.2018.12.059
|
| [41] | Sterl S, Vanderkelen I, Chawanda CJ, et al. (2020) Smart renewable electricity portfolios in West Africa. Nat Sustain |
| [42] |
Oyewo AS, Aghahosseini A, Ram M, et al. (2020) Transition towards decarbonized power systems and its socio-economic impacts in West Africa. Renewable Energy 154: 1092-1112. doi: 10.1016/j.renene.2020.03.085
|
| [43] |
Jurasz J, Beluco A, Canales FA (2018) The impact of complementarity on power supply reliability of small scale hybrid energy systems. Energy 161: 737-743. doi: 10.1016/j.energy.2018.07.182
|
| [44] |
Demissie AA, Solomon AA (2016) Power system sensitivity to extreme hydrological conditions as studied using an integrated reservoir and power system dispatch model, the case of Ethiopia. Appl Energ 182: 442-463. doi: 10.1016/j.apenergy.2016.08.106
|
| [45] |
Barbosa LDSNS, Orozco JF, Bogdanov D, et al. (2016) Hydropower and power-to-gas storage options: The Brazilian energy system case. Energy Procedia 99: 89-107. doi: 10.1016/j.egypro.2016.10.101
|
| [46] |
Solomon AA, Child M, Caldera U, et al. (2017) How much energy storage is needed to incorporate very large intermittent renewables? Energy Procedia 135: 283-293. doi: 10.1016/j.egypro.2017.09.520
|
| [47] |
Pensini A, Rasmussen CN, Kempton W (2014) Economic analysis of using excess renewable electricity to displace heating fuels. Appl Energ 131: 530-543. doi: 10.1016/j.apenergy.2014.04.111
|
| [48] | Mensah TNO The role of modern bioenergy in solar photovoltaic driven and defossilised power systems-The case of Ghana. MSc Thesis, LUT University, Lappeenranta, 2019 |
| [49] |
Weschenfelder F, Leite GNP, Costa AC, et al. (2020) A review on the complementarity between grid-connected solar and wind power systems. J Clean Prod 257: 120617. doi: 10.1016/j.jclepro.2020.120617
|
| [50] |
Jurasz J, Canales FA, Kies A, et al. (2020) A review on the complementarity of renewable energy sources: Concept, metrics, application and future research directions. Sol Energy 195: 703-724. doi: 10.1016/j.solener.2019.11.087
|
| [51] | Yan J, Qu T, Han S, et al. (2020) Reviews on characteristic of renewables: Evaluating the variability and complementarity. Int T Electr Energy 30: e12281. |
| [52] |
Liu LJ, Wang Z, Wang Y, et al. (2020) Optimizing wind/solar combinations at finer scales to mitigate renewable energy variability in China. Renewable Sustainable Energy Rev 132: 110151. doi: 10.1016/j.rser.2020.110151
|
| [53] |
Xu L, Wang ZW, Liu YF (2017) The spatial and temporal variation features of wind-sun complementarity in China. Energ Convers Manage 154: 138-148. doi: 10.1016/j.enconman.2017.10.031
|
| [54] |
Zhang HX, Cao YJ, Zhang Y, et al. (2018) Quantitative synergy assessment of regional wind-solar energy resources based on MERRA reanalysis data. Appl Energ 216: 172-182. doi: 10.1016/j.apenergy.2018.02.094
|
| [55] | Holttinen H, Kiviluoma J, Levy T, et al. (2016) Design and Operation of Power Systems with Large Amounts of Wind Power: Final Summary Report, IEA WIND Task 25. Available from: https://www.vttresearch.com/sites/default/files/pdf/technology/2016/T268.pdf. |
| [56] |
Child M, Kemfert C, Bogdanov D, et al. (2019) Flexible electricity generation, grid exchange and storage for the transition to a 100% renewable energy system in Europe. Renewable Energy 139: 80-101. doi: 10.1016/j.renene.2019.02.077
|
| [57] | Cui M, Zhang J, Florita AR, et al. (2015) An Optimized swinging door algorithm for wind power ramp event detection. 2015 IEEE Power & Energy Society General Meeting, 26-30. |
| [58] |
Debnath K, Goel L (1995) Power system planning-a reliability perspective. Electr Pow Syst Res 34: 179-185. doi: 10.1016/0378-7796(95)00976-X
|
| [59] | Breyer C, Bogdanov D, Aghahosseini A, et al. (2020) On the techno-economic benefits of a global energy interconnection. EEEP 9: 83-102. |
| [60] |
Bogdanov D, Farfan J, Sadovskaia K, et al. (2019) Radical transformation pathway towards sustainable electricity via evolutionary steps. Nat Commun 10: 1077. doi: 10.1038/s41467-019-08855-1
|
| [61] | Ram M, Bogdanov D, Aghahosseini A, et al. (2019) Global Energy System Based on 100% Renewable Energy-Power, Heat, Transport and Desalination Sectors, Lappeenranta: LUT University and Energy Watch Group. |
| [62] | Ram M, Bogdanov D, Aghahosseini A, et al. (2020) 100% Renewable Europe: How to Make Europe's Energy System Climate-Neutral Before 2050, Brussels and Lappeenranta: SolarPower Europe and LUT University. |
| [63] |
Caldera U, Bogdanov D, Breyer C (2016) Local cost of seawater RO desalination based on solar PV and wind energy-A global estimate. Desalination 385: 207-216. doi: 10.1016/j.desal.2016.02.004
|
| [64] |
Breyer C, Bogdanov D, Gulagi A, et al. (2017) On the role of solar photovoltaics in global energy transition scenarios. Prog Photovolt 25: 727-745. doi: 10.1002/pip.2885
|
| [65] |
Soder L, Tomasson E, Estanqueiro A, et al. (2020) Review of wind generation within adequacy calculations and capacity markets for different power systems. Renewable Sustainable Energy Rev 119: 109540. doi: 10.1016/j.rser.2019.109540
|
| [66] |
Caldera U, Breyer C (2018) The role that battery and water storage play in Saudi Arabia's transition to an integrated 100% renewable energy power system. J Energy Storage 17: 299-310. doi: 10.1016/j.est.2018.03.009
|
| [67] |
Azzuni A, Breyer C (2020) Global Energy Security index and its Application on National level. Energies 13: 2502. doi: 10.3390/en13102502
|
| [68] |
Breyer C, Tsupari E, Tikka V, et al. (2015) Power-to-gas as an emerging profitable business through creating an integrated value chain. Energy Procedia 73: 182-189. doi: 10.1016/j.egypro.2015.07.668
|
| [69] |
Child M, Nordling A, Breyer C (2018) The impacts of high V2G participation in a 100% renewable Åland energy system. Energies 11: 2206. doi: 10.3390/en11092206
|
| [70] | Taljegard M, Walter V, Goransson L, et al. (2019) Impact of electric vehicles on the cost-competitiveness of generation and storage technologies in electricity system. Environ Res Lett 14: 12. |
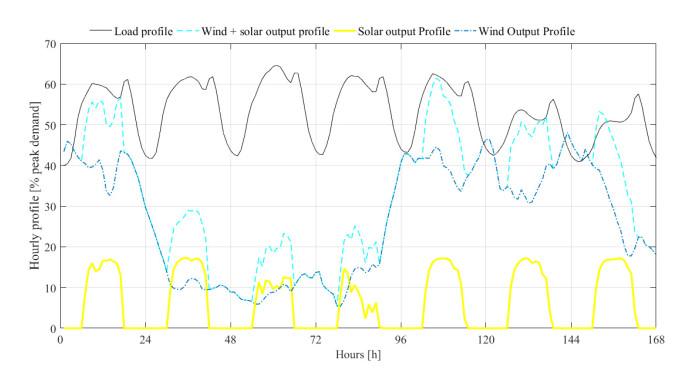
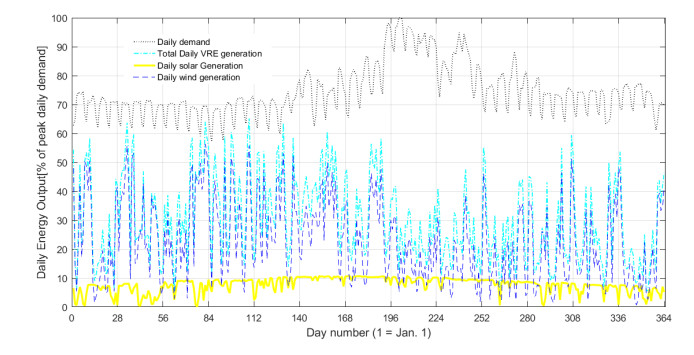
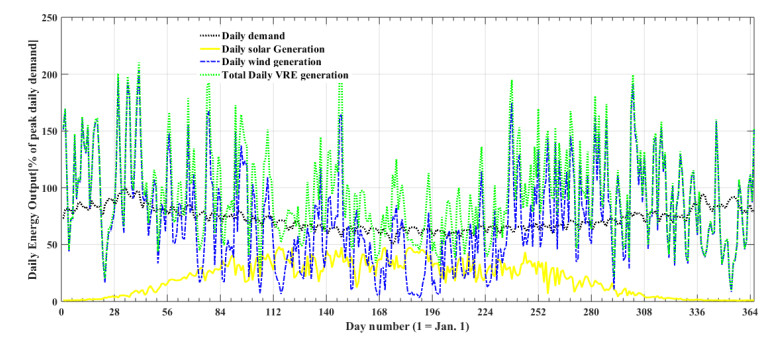
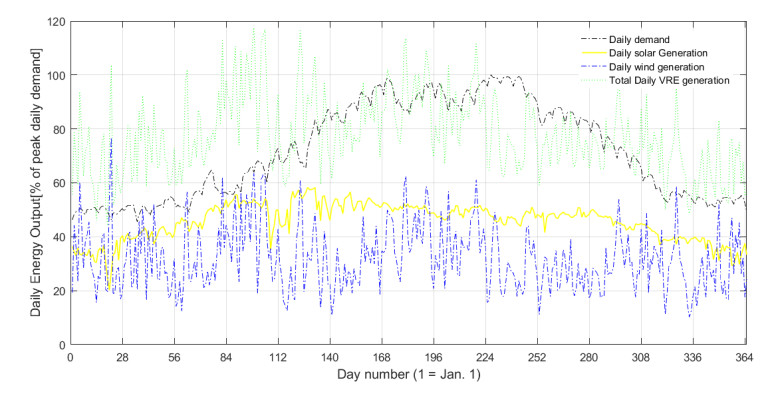
Figures(5) / Tables(5)

Solomon A. A., Michel Child, Upeksha Caldera, Christian Breyer. Exploiting wind-solar resource complementarity to reduce energy storage need[J]. AIMS Energy, 2020, 8(5): 749-770. doi: 10.3934/energy.2020.5.749







 DownLoad:
DownLoad: