Citation: Maria Jose Miguez, Wenyaw Chan, Luis Espinoza, Ralph Tarter, Caroline Perez. Marijuana use among adolescents is associated with deleterious alterations in mature BDNF[J]. AIMS Public Health, 2019, 6(1): 4-14. doi: 10.3934/publichealth.2019.1.4

| [1] | World Health Organization. Substance Abuse: Facts and Figures, 2015. Available from: http://www.who.int/substance_abuse/facts/cannabis/en/. |
| [2] |
Jacobus J, Tapert SF (2014) Effects of Cannabis on the Adolescent Brain. Curr Pharm Des 20: 2186–2193. doi: 10.2174/13816128113199990426

|
| [3] | World Drug Report 2014. United Nations publication, Sales No. E.14.XI.7. Available from: http://www.unodc.org/documents/wdr2014/World_Drug_Report_2014_web.pdf |
| [4] |
Sevigny EL (2013) Is today's marijuana more potent simply because it's fresher? Drug Test Anal 5: 62–67. doi: 10.1002/dta.1430
|
| [5] |
D'Souza DC, Pittman B, Perry E (2009) Simen A Preliminary evidence of cannabinoid effects on brain-derived neurotrophic factor (BDNF) levels in humans. Psychopharmacology 202: 569. doi: 10.1007/s00213-008-1333-2
|
| [6] |
Jockers-Scherübl MC, Danker-Hopfe H, Mahlberg R, et al. (2004) Brain-derived neurotrophic factor serum concentrations are increased in drug-naive schizophrenic patients with chronic cannabis abuse and multiple substance abuse. Neurosci Lett 371: 79–83. doi: 10.1016/j.neulet.2004.08.045
|
| [7] | Arseneault L, Cannon M, Poulton R, et al. (2002) Cannabis use in adolescence and risk for adult psychosis: longitudinal prospective study. BMJ 7374: 1212–1213. |
| [8] |
Karege F, Perret G, Bondolfi G, et al. (2002) Decreased serum brain-derived neurotrophic factor levels in major depressed patients. Psychiatry Res 109: 143–148. doi: 10.1016/S0165-1781(02)00005-7
|
| [9] | NIDA Marijuana National Institute on Drug Abuse, 2017. Available from: https://www.drugabuse.gov/publications/research-reports/marijuana. |
| [10] |
Freedland CS, Whitlow CT, Miller MD, et al. (2002) Dose‐dependent effects of Δ9‐tetrahydrocannabinol on rates of local cerebral glucose utilization in rat. Synapse 45: 134–142. doi: 10.1002/syn.10089
|
| [11] |
Atakan Z (2012) Cannabis, a complex plant: different compounds and different effects on individuals. Ther Adv Psychopharmacol 2: 241–254. doi: 10.1177/2045125312457586
|
| [12] |
Berghuis P, Dobszay MB, Wang X, et al. (2005) Endocannabinoids regulate interneuron migration and morphogenesis by transactivating the TrkB receptor. Proc Natl Acad Sci U S A 102: 19115–19120. doi: 10.1073/pnas.0509494102
|
| [13] |
Angelucci F, Ricci V, Spalletta G, et al. (2008) Reduced serum concentrations of nerve growth factor, but not brain-derived neurotrophic factor, in chronic cannabis abusers. Eur Neuropsychopharmacol 18: 882–887. doi: 10.1016/j.euroneuro.2008.07.008
|
| [14] | Klug M, Maarten VDB (2013) An investigation into "two hit" effects of BDNF deficiency and young-adult cannabinoid receptor stimulation on prepulse inhibition regulation and memory in mice. Front Behav Neurosci 7: 149. |
| [15] |
Boulle F, Van Den Hove DL, Jakob SB et al. (2012) Epigenetic regulation of the BDNF gene: implications for psychiatric disorders. Mol Psychiatry 17: 584. doi: 10.1038/mp.2011.107
|
| [16] |
Jones S, Bonci A (2005) Synaptic plasticity and drug addiction. Curr Opin Pharmacol 5: 20–25. doi: 10.1016/j.coph.2004.08.011
|
| [17] |
Jacobus J, Tapert SF (2014) Effects of Cannabis on the Adolescent Brain. Curr Pharm Des 20: 2186–2193. doi: 10.2174/13816128113199990426
|
| [18] | Weiland BJ, Thayer RE, Depue BE, et al. (2015) Daily marijuana use is not associated with brain morphometric measures in adolescents or adults. J Neurosci 35: 1505–1512. |
| [19] |
Gilman JM, Kuster JK, Lee S, et al. (2014) Cannabis Use Is Quantitatively Associated with Nucleus Accumbens and Amygdala Abnormalities in Young Adult Recreational Users. J Neurosci 34: 5529–5538. doi: 10.1523/JNEUROSCI.4745-13.2014
|
| [20] | National Institute on Drug Abuse. The NIDA Quick Screen. Screening for Drug Use in General Medical Settings: Resource Guide, March, 2012. Available from: https://www.drugabuse.gov/drugs-abuse/opioids |
| [21] |
Levy S, Weiss R, Sherritt L, et al. (2014) An Electronic Screen for Triaging Adolescent Substance Use by Risk Levels. JAMA Pediatr 168: 822–828. doi: 10.1001/jamapediatrics.2014.774
|
| [22] |
Lu B (2003) Pro-region of neurotrophins: role in synaptic modulation. Neuron 39: 735–738. doi: 10.1016/S0896-6273(03)00538-5
|
| [23] |
Klein AB, Williamson R, Santini MA, et al. (2011) Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int J Neuropsychopharmacol 14: 347–353. doi: 10.1017/S1461145710000738
|
| [24] |
Richardson MT, Ainsworth BE, Jacobs DR, et al. (2001) Validation of the Stanford 7-day recall to assess habitual physical activity. Ann Epidemiol 11: 145–153. doi: 10.1016/S1047-2797(00)00190-3
|
| [25] |
Koh HK, Blakey CR, Roper AY (2014) Healthy People 2020: a report card on the health of the nation. JAMA 311: 2475–2476. doi: 10.1001/jama.2014.6446
|
| [26] |
Konings M, Henquet C, Maharajh HD, et al. (2008) Early exposure to cannabis and risk for psychosis in young adolescents in Trinidad. Acta Psychiatr Scand 118: 209–213. doi: 10.1111/j.1600-0447.2008.01202.x
|
| [27] | Jessor R, Chase JA, Donovan JE (1980) Psychosocial correlates of marijuana use and problem drinking in a national sample of adolescents. Am J Public Health 6: 604–613. |
| [28] |
Stefanis NC, Delespaul P, Henquet C, et al. (2004) Early adolescent cannabis exposure and positive and negative dimensions of psychosis. Addiction 99: 1333–1341. doi: 10.1111/j.1360-0443.2004.00806.x
|
| [29] |
Decoster J, van Os J, Kenis G, et al. (2011) Age at onset of psychotic disorder: Cannabis, BDNF Val66Met, and sex‐specific models of gene–environment interaction. Am J Med Genet B Neuropsychiatr Genet 156: 363–369. doi: 10.1002/ajmg.b.31174
|
| [30] | Hill AB (1965) The Environment and Disease: Association or Causation? Proc R Soc Med 58: 295–300. |
| [31] |
Yücel M, Lorenzetti V, Suo C, et al. (2016) Hippocampal harms, protection and recovery following regular cannabis use. Transl Psychiatry 6: e710. doi: 10.1038/tp.2015.201
|
| [32] | Shieh PB, Ghosh A (1999) Molecular mechanisms underlying activity-dependent regulation of BDNF expression. J Neurobiol 1: 127–134. |
| [33] |
Butovsky E, Juknat A, Goncharov I, et al. (2005) In vivo up‐regulation of brain‐derived neurotrophic factor in specific brain areas by chronic exposure to Δ9‐tetrahydrocannabinol. J neurochemistry 93: 802–811. doi: 10.1111/j.1471-4159.2005.03074.x
|
| [34] | Murray PS, Holmes PV (2011) An Overview of Brain-Derived Neurotrophic Factor and Implications for Excitotoxic Vulnerability in the Hippocampus. Int J Pept 654085. |
| [35] |
Monteggia LM, Barrot M, Powell CM, et al. (2004) Essential role of brain-derived neurotrophic factor in adult hippocampal function. Proc Natl Acad Sci U S A 101: 10827–10832. doi: 10.1073/pnas.0402141101
|
| [36] |
Kleim JA, Chan S, Pringle E, et al. (2006) BDNF val66met polymorphism is associated with modified experience-dependent plasticity in human motor cortex. Nat Neurosci 9: 735–737. doi: 10.1038/nn1699
|
| [37] |
Beste C, Kolev V, Yordanova J, et al. (2010) The role of the BDNF Val66Met polymorphism for the synchronization of error-specific neural networks. J Neurosci 30: 10727–10733. doi: 10.1523/JNEUROSCI.2493-10.2010
|
| [38] |
Li X, Wolf ME (2015) Multiple faces of BDNF in cocaine addiction. Behav Brain Res 279: 240–254. doi: 10.1016/j.bbr.2014.11.018
|
| [39] |
Vargas-Perez H, Ting-A Kee R, Walton CH, et al. (2009) Ventral tegmental area BDNF induces an opiate-dependent-like reward state in naïve rats. Science 324:1732–1734. doi: 10.1126/science.1168501
|
| [40] |
Verheij MM, Vendruscolo LF, Caffino L, et al. (2016) Systemic delivery of a brain-penetrant TrkB antagonist reduces cocaine self-administration and normalizes TrkB signaling in the nucleus accumbens and prefrontal cortex. J Neurosci 36: 8149–8159. doi: 10.1523/JNEUROSCI.2711-14.2016
|
| [41] | McGinty JF, Whitfield TW, Berglind WJ (2010) Brain-derived neurotrophic factor and cocaine addiction. Brain Res 1314C: 183. |
| [42] |
Fujimura H, Altar CA, Chen R, et al. (2002) Brain-derived neurotrophic factor is stored in human platelets and released by agonist stimulation. Thromb Haemost 87: 728–734. doi: 10.1055/s-0037-1613072
|
| [43] |
Pan W, Banks WA, Fasold MB, et al. (1998) Transport of brain-derived neurotrophic factor across the blood-brain barrier. Neuropharmacology 37: 1553–1561. doi: 10.1016/S0028-3908(98)00141-5
|
| [44] |
Radka SF, Holst PA, Fritsche M, et al. (1996) Presence of brain-derived neurotrophic factor in brain and human and rat but not mouse serum detected by a sensitive and specific immunoassay. Brain Res 709: 122–301. doi: 10.1016/0006-8993(95)01321-0
|
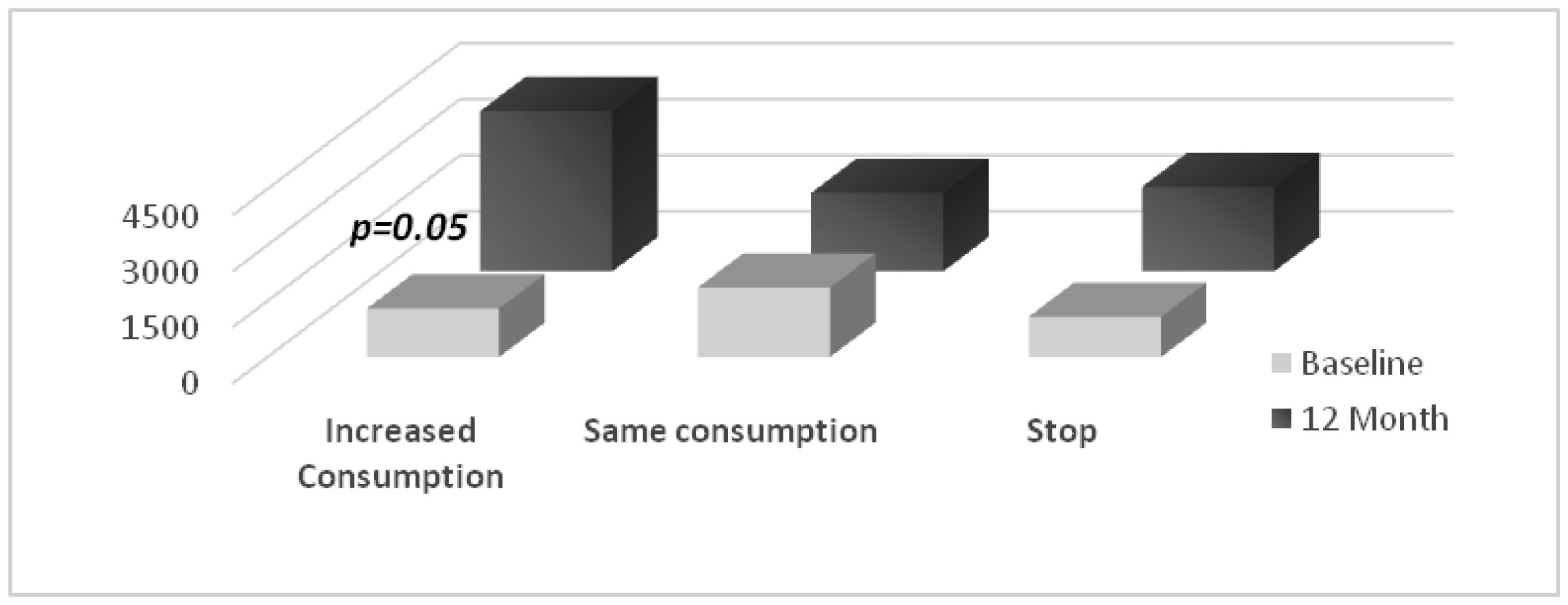
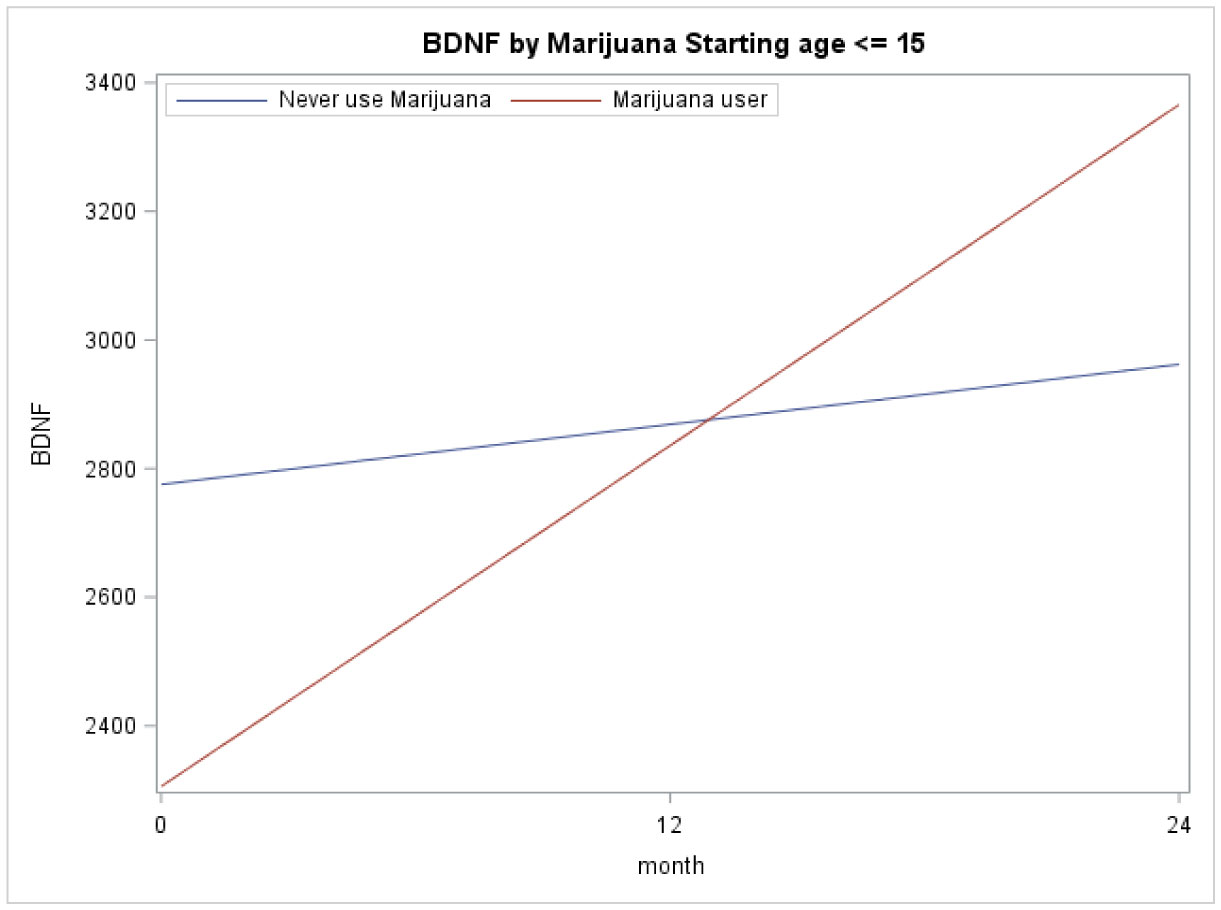
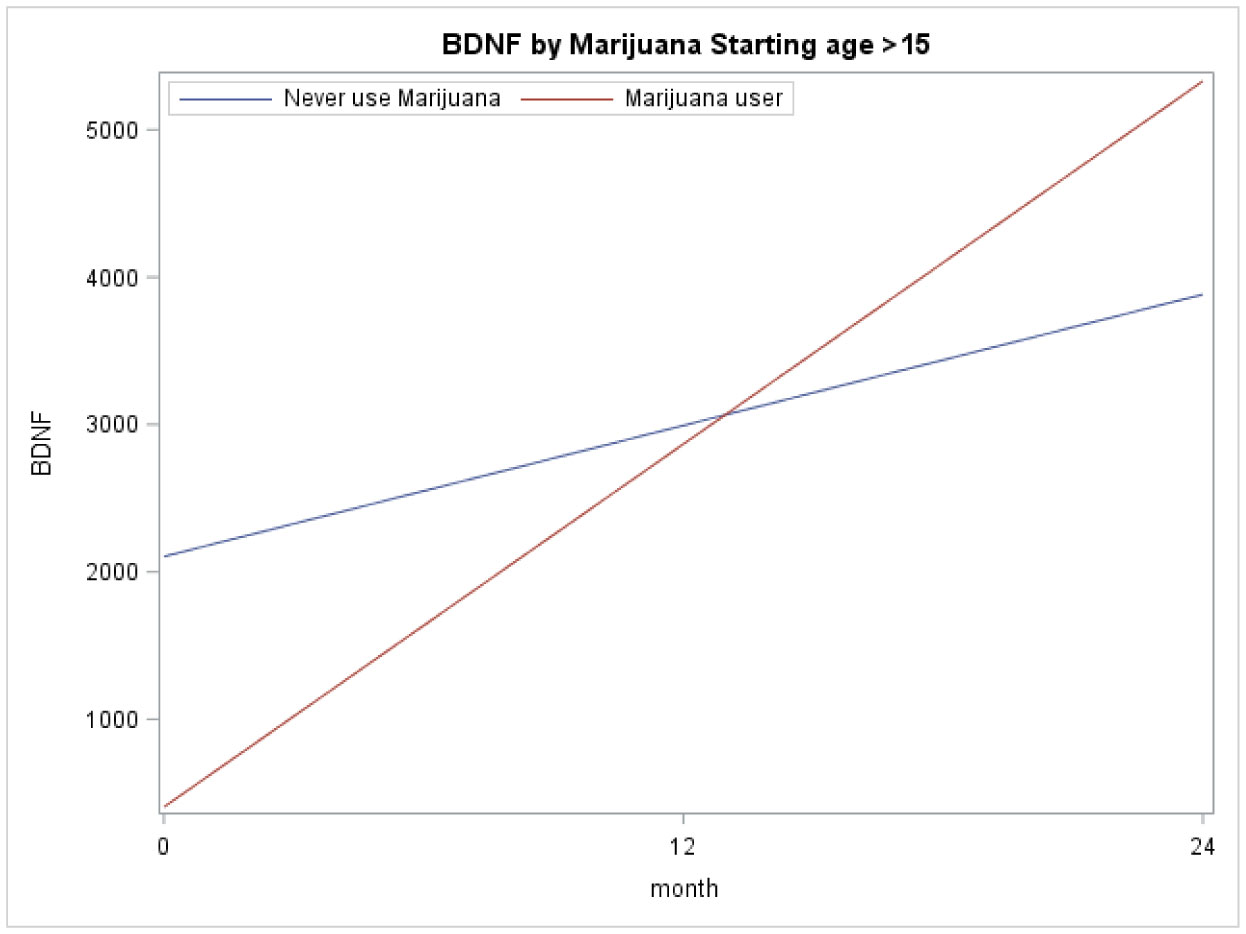
Figures(3) / Tables(2)

Maria Jose Miguez, Wenyaw Chan, Luis Espinoza, Ralph Tarter, Caroline Perez. Marijuana use among adolescents is associated with deleterious alterations in mature BDNF[J]. AIMS Public Health, 2019, 6(1): 4-14. doi: 10.3934/publichealth.2019.1.4







 DownLoad:
DownLoad: