Citation: Yingyu Law, Leo Wein. Reversing the nutrient drain through urban insect farming—opportunities and challenges[J]. AIMS Bioengineering, 2018, 5(4): 226-237. doi: 10.3934/bioeng.2018.4.226

| [1] | Alexandratos N, Bruinsma J (2012) World agriculture towards 2030/2050: The 2012 revision. ESA Working Paper No. 12-03, Food and Agriculture Organization of the United Nations (FAO) , Rome. |
| [2] |
Godfray HCJ, Beddington JR, Crute IR, et al. (2010) Food security: The challenge of feeding 9 billion people. Science 327: 812–818. doi: 10.1126/science.1185383

|
| [3] | Oliva-Teles A, Enes P, Peres H (2015) Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish, In: Davis DA, Feed and Feeding Practices in Aquaculture, 8 Eds., Oxford: Woodhead Publishing, 203–233. |
| [4] | Skøt J, Lipper L, Thomas G, et al. (2016) The state of food and agriculture: Climate change, agriculture and food security. Food and Agriculture Organization of the United Nations (FAO), Rome. |
| [5] | Food and Agriculture Organization of the United Nations (FAO) (2013) Food wastage footprint: Impact on natural resources. Summary report. Rome. |
| [6] |
Tilman D, Fargione J, Wolff B, et al. (2001) Forecasting agriculturally driven global environmental change. Science 292: 281–284. doi: 10.1126/science.1057544
|
| [7] |
Zhao C, Liu B, Piao S, et al. (2017) Temperature increase reduces global yields of major crops in four independent estimates. P Natl Acad Sci USA 114: 9326–9331. doi: 10.1073/pnas.1701762114
|
| [8] |
Adhikari B, Barrington S, Martinez J (2009) Urban food waste generation: Challenges and opportunities. Int J Environ Waste Manage 3: 4–21. doi: 10.1504/IJEWM.2009.024696
|
| [9] |
Hoornweg D, Bhada-Tata P, Kennedy C (2013) Environment: Waste production must peak this century. Nature 502: 615–617. doi: 10.1038/502615a
|
| [10] | Gustavsson J, Cederberg C, Sonesson U, et al. (2011) Global fodd losses and food waste. extent, causes and prevention. Food and Agriculture Organization of the United Nations (FAO), Rome, Italy. |
| [11] |
Diener S, Zurbrügg C, Tockner K (2009) Conversion of organic material by black soldier fly larvae: Establishing optimal feeding rates. Waste Manag Res 27: 603–610. doi: 10.1177/0734242X09103838
|
| [12] |
St-Hilaire S, Sheppard C, Tomberlin JK, et al. (2007) Fly Prepupae as a Feedstuff for Rainbow Trout, Oncorhynchus mykiss. J World Aquacul Soc 38: 59–67. doi: 10.1111/j.1749-7345.2006.00073.x
|
| [13] |
Newton GL, Booram CV, Barker RW, et al. (1977) Dried Hermetia illucens larvae meal as a supplement for swine. J Anim Sci 44: 395–400. doi: 10.2527/jas1977.443395x
|
| [14] |
Lee J, Kim YM, Park YK, et al. (2018) Black soldier fly (Hermetia illucens) larvae enhances immune activities and increases survivability of broiler chicks against experimental infection of Salmonella Gallinarum. J Vet Med Sci 80: 736–740. doi: 10.1292/jvms.17-0236
|
| [15] | Kupferschmidt K (2015) Feature: Why insects could be the ideal animal feed. Science. |
| [16] | Alltech (2017) 7th Annual Alltech Global Feed Survey 2018. Available from: https://go.alltech.com/alltech-feed-survey. |
| [17] | Gilbert R (2004) The world animal feed industry. Protein Sources for the Animal Feed Industry. Food and Agriculture Organization of the United Nations (FAO). Rome, Italy. |
| [18] | SEAFISH (2016) Fishmeal and fish oil facts and figures. Available from: http://www.seafish.org/media/Publications/SeafishFishmealandFishOilFactsandFigures_201612.pdf. |
| [19] | Food and Agriculture Organization of the United Nations (FAO) (2018)The State of World Fisheries and Aquaculture: Meeting the sustainable development goals. Rome. |
| [20] | Masuda T, Goldsmith PD (2009) World soybean production: Area harvested, yield, and long-term projections. Int Food Agribus Man 12: 143–163. |
| [21] | Department of Agriculture (US) (2018) Fish Meal Production by Country in 1000 MT. Available from: https://www.indexmundi.com/agriculture/?commodity=fish-meal: Index Mundi. |
| [22] |
Tacon AGJ, Metian M (2008) Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 285: 146–158. doi: 10.1016/j.aquaculture.2008.08.015
|
| [23] | Trostle R (2010) Global Agricultural Supply and Demand: Factors Contributing to the Recent Increase in Food Commodity Prices. Available from: https://www.hsdl.org/?abstract&did=485652. |
| [24] |
Verkerk MC, Tramper J, van Trijp JCM, et al. (2007) Insect cells for human food. Biotechnol Adv 25: 198–202. doi: 10.1016/j.biotechadv.2006.11.004
|
| [25] |
Dobermann D, Swift JA, Field LM (2017) Opportunities and hurdles of edible insects for food and feed. Nutr Bull 42: 293–308. doi: 10.1111/nbu.12291
|
| [26] |
van Huis A (2013) Potential of insects as food and feed in assuring food security. Annu Rev Entomol 58: 563–583. doi: 10.1146/annurev-ento-120811-153704
|
| [27] | Hilkens W, de Klerk B (2017) The cultivation of insects: A small sector with great opportunities. Netherlands: ABN AMRO and BOM. |
| [28] |
Tomberlin JK, Sheppard DC, Joyce JA (2002) Selected Life-History Traits of Black Soldier Flies (Diptera: Stratiomyidae) Reared on Three Artificial Diets. Ann Entomol Soc Am 95: 379–386. doi: 10.1603/0013-8746(2002)095[0379:SLHTOB]2.0.CO;2
|
| [29] |
Sheppard C (1983) House Fly and Lesser Fly Control Utilizing the Black Soldier Fly in Manure Management Systems for Caged Laying Hens. Environ Entomol 12: 1439–1442. doi: 10.1093/ee/12.5.1439
|
| [30] |
Čičková H, Newton GL, Lacy RC, et al. (2015) The use of fly larvae for organic waste treatment. Waste Manage 35: 68–80. doi: 10.1016/j.wasman.2014.09.026
|
| [31] |
Nguyen TTX, Tomberlin JK, Vanlaerhoven S (2015) Ability of Black Soldier Fly (Diptera: Stratiomyidae) larvae to recycle food waste. Environ Entomol 44: 406–410. doi: 10.1093/ee/nvv002
|
| [32] |
Li W, Li Q, Zheng L, et al. (2015) Potential biodiesel and biogas production from corncob by anaerobic fermentation and black soldier fly. Bioresource Technol 194: 276–282. doi: 10.1016/j.biortech.2015.06.112
|
| [33] | Spranghers T, Ottoboni M, Klootwijk C, et al. (2016) Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J Sci Food Agr 97: 2594–2600. |
| [34] | Cammack AJ, Tomberlin KJ (2017) The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.) (Diptera: Stratiomyidae). Insects 8: 56. |
| [35] | Cheng JYK, Chiu SLH, Lo IMC (2017) Effects of moisture content of food waste on residue separation, larval growth and larval survival in black soldier fly bioconversion. Waste Manage 67: 315–323. |
| [36] |
Lardé G (1989) Investigation on some factors affecting larval growth in a coffee-pulp bed. Biol Wastes 30: 11–19. doi: 10.1016/0269-7483(89)90139-0
|
| [37] |
Gupta S, Lee JJL, Chen WN (2018) Analysis of improved nutritional composition of potential functional food (Okara) after probiotic solid-state fermentation. J Agr Food Chem 66: 5373–5381. doi: 10.1021/acs.jafc.8b00971
|
| [38] |
Jucker C, Erba D, Leonardi MG, et al. (2017) Assessment of vegetable and fruit substrates as potential rearing media for Hermetia illucens (diptera: stratiomyidae) larvae. Environ Entomol 46: 1415–1423. doi: 10.1093/ee/nvx154
|
| [39] | Rehman Ku, Rehman A, Cai M, et al. (2017) Conversion of mixtures of dairy manure and soybean curd residue by black soldier fly larvae (Hermetia illucens L.). J Clean Prod 154: 366–373. |
| [40] |
Tschirner M, Simon A (2015) Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J Insects Food Feed 1: 249–259. doi: 10.3920/JIFF2014.0008
|
| [41] |
Ma J, Lei Y, Rehman KU, et al. (2018) Dynamic effects of initial pH of substrate on biological growth and metamorphosis of black soldier fly (Diptera: Stratiomyidae). Environ Entomol 47: 159–165. doi: 10.1093/ee/nvx186
|
| [42] |
Paz ASP, Carrejo NS, Rodríguez CHG (2015) Effects of larval density and feeding rates on the bioconversion of vegetable waste using black soldier fly larvae Hermetia illucens (L.), (Diptera: Stratiomyidae). Waste Biomass Valorization 6: 1059–1065. doi: 10.1007/s12649-015-9418-8
|
| [43] |
Harnden LM, Tomberlin JK (2016) Effects of temperature and diet on black soldier fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), development. Forensic Sci Int 266: 109–116. doi: 10.1016/j.forsciint.2016.05.007
|
| [44] |
Liu X, Chen X, Wang H, et al. (2017) Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS One 12: e0182601. doi: 10.1371/journal.pone.0182601
|
| [45] | Dortmans B, Diener S, Verstappen B, et al. (2017) Black soldier fly biowaste processing. A step-by step guide. |
| [46] | Kok R (2017) Insect Production and Facility Design, In: Van Huis A, Tomberlin JK, Insects as food and feed: From production to consumption, 8 Eds., Wageningen: Wageningen Academic Publishers, 143–172. |
| [47] |
Basset Y, Cizek L, Cuénoud P, et al. (2012) Arthropod diversity in a tropical forest. Science 338: 1481–1484. doi: 10.1126/science.1226727
|
| [48] |
Engel P, Moran NA (2013) The gut microbiota of insects-diversity in structure and function. FEMS Microbiol Rev 37: 699–735. doi: 10.1111/1574-6976.12025
|
| [49] |
The Human Microbiome Project C (2012) Structure, function and diversity of the healthy human microbiome. Nature 486: 207–214. doi: 10.1038/nature11234
|
| [50] |
Chaucheyras-Durand F, Durand H (2010) Probiotics in animal nutrition and health. Benefic Microbes 1: 3–9. doi: 10.3920/BM2008.1002
|
| [51] | Zheng L, Crippen TL, Holmes L, et al. (2013) Bacteria mediate oviposition by the black soldier fly, Hermetia illucens (L.), (Diptera: Stratiomyidae). Sci Rep 3: 2563. |
| [52] |
Chandler JA, Lang JM, Bhatnagar S, et al. (2011) Bacterial communities of diverse drosophila species: Ecological context of a host–microbe model system. PLoS Genet 7: e1002272. doi: 10.1371/journal.pgen.1002272
|
| [53] | Klammsteiner T, Walter A, Heussler C, et al. (2018) Hermetia illucens (Diptera: Stratiomyidae) larvae in waste valorization and diet-based shifts in their gut microbiome. Available from: http://uest.ntua.gr/naxos2018/proceedings/pdf/86_NAXOS2018_Klammsteiner_etal.pdf. |
| [54] |
Vogel H, Muller A, Heckel DG, et al. (2018) Nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev Comp Immunol 78: 141–148. doi: 10.1016/j.dci.2017.09.008
|
| [55] |
Erickson MC, Islam M, Sheppard C, et al. (2004) Reduction of Escherichia coli O157:H7 and Salmonella enterica serovar Enteritidis in chicken manure by larvae of the black soldier fly. J Food Prot 67: 685–690. doi: 10.4315/0362-028X-67.4.685
|
| [56] |
Liu Q, Tomberlin JK, Brady JA, et al. (2008) Black soldier fly (Diptera: Stratiomyidae) larvae reduce Escherichia coli in dairy manure. Environ Entomol 37: 1525–1530. doi: 10.1603/0046-225X-37.6.1525
|
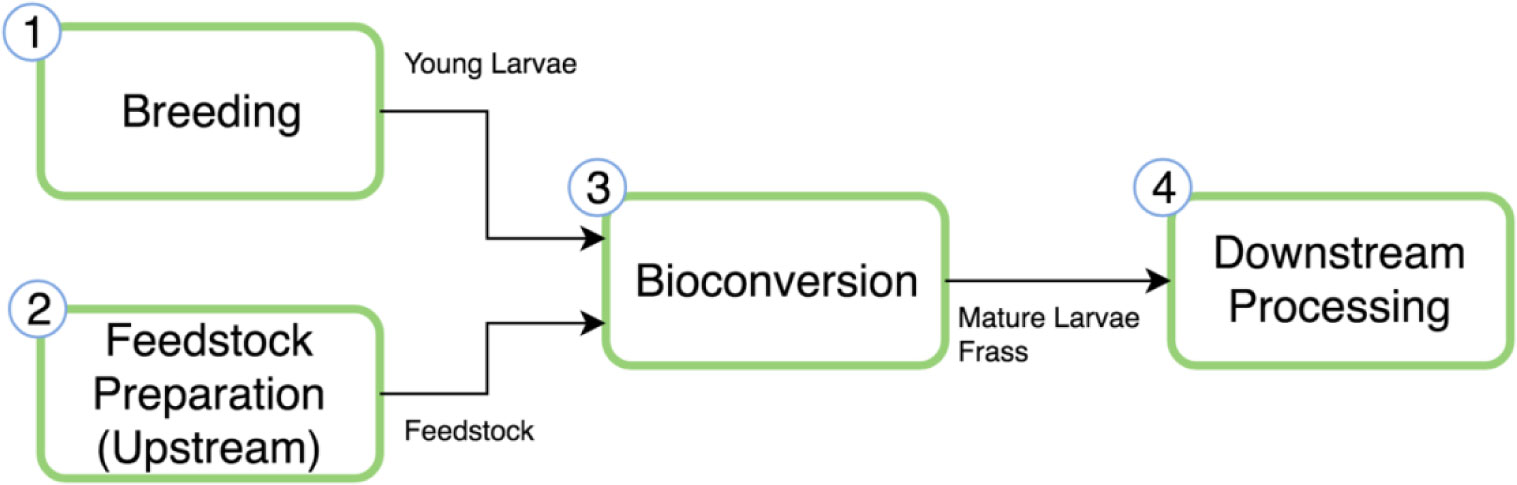
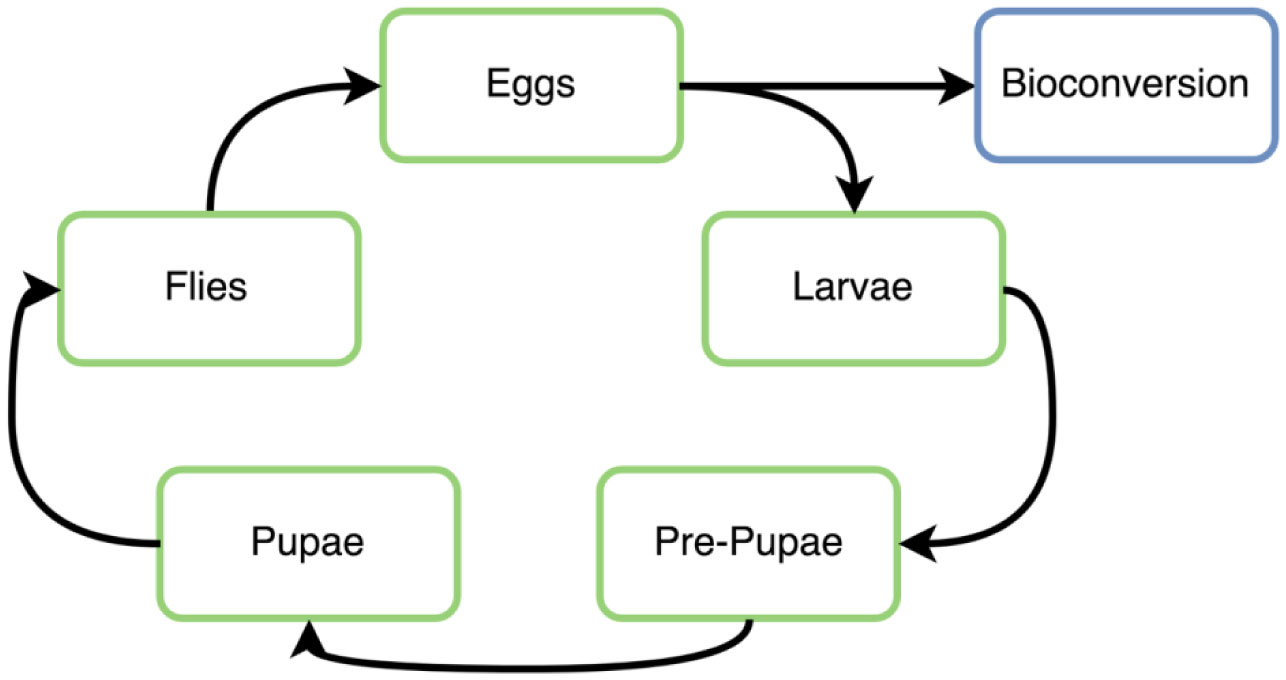
Figures(2)

Yingyu Law, Leo Wein. Reversing the nutrient drain through urban insect farming—opportunities and challenges[J]. AIMS Bioengineering, 2018, 5(4): 226-237. doi: 10.3934/bioeng.2018.4.226







 DownLoad:
DownLoad: