Citation: Sebastian Kube, Petra Wendler. Structural comparison of contractile nanomachines[J]. AIMS Biophysics, 2015, 2(2): 88-115. doi: 10.3934/biophy.2015.2.88

| [1] | Leiman PG, Shneider MM (2012) Contractile tail machines of bacteriophages. In: Rossmann MG, Rao VB, editors. Viral Molecular Machines. New York Dordrecht Heidelberg London: Springer. 93-114. |
| [2] |
Michel-Briand Y, Baysse C (2002) The pyocins of Pseudomonas aeruginosa. Biochimie 84: 499-510. doi: 10.1016/S0300-9084(02)01422-0

|
| [3] |
Sarris PF, Ladoukakis ED, Panopoulos NJ, et al. (2014) A phage tail-derived element with wide distribution among both prokaryotic domains: a comparative genomic and phylogenetic study. Genome Biol Evol 6: 1739-1747. doi: 10.1093/gbe/evu136
|
| [4] |
Amos LA, Klug A (1975) Three-dimensional image reconstructions of the contractile tail of T4 bacteriophage. J Mol Biol 99: 51-64. doi: 10.1016/S0022-2836(75)80158-6
|
| [5] |
Leiman PG, Chipman PR, Kostyuchenko VA, et al. (2004) Three-dimensional rearrangement of proteins in the tail of bacteriophage T4 on infection of its host. Cell 118: 419-429. doi: 10.1016/j.cell.2004.07.022
|
| [6] |
Kostyuchenko VA, Chipman PR, Leiman PG, et al. (2005) The tail structure of bacteriophage T4 and its mechanism of contraction. Nat Struct Mol Biol 12: 810-813. doi: 10.1038/nsmb975
|
| [7] |
Effantin G, Hamasaki R, Kawasaki T, et al. (2013) Cryo-electron microscopy three-dimensional structure of the jumbo phage PhiRSL1 infecting the phytopathogen Ralstonia solanacearum. Structure 21: 298-305. doi: 10.1016/j.str.2012.12.017
|
| [8] |
Fokine A, Battisti AJ, Bowman VD, et al. (2007) Cryo-EM study of the Pseudomonas bacteriophage phiKZ. Structure 15: 1099-1104. doi: 10.1016/j.str.2007.07.008
|
| [9] |
Aksyuk AA, Kurochkina LP, Fokine A, et al. (2011) Structural conservation of the myoviridae phage tail sheath protein fold. Structure 19: 1885-1894. doi: 10.1016/j.str.2011.09.012
|
| [10] |
Schwarzer D, Buettner FF, Browning C, et al. (2012) A multivalent adsorption apparatus explains the broad host range of phage phi92: a comprehensive genomic and structural analysis. J Virol 86: 10384-10398. doi: 10.1128/JVI.00801-12
|
| [11] | Ge P, Scholl D, Leiman PG, et al. (2015) Atomic structures of a bactericidal contractile nanotube in its pre- and postcontraction states. Nat Struct Mol Biol. |
| [12] |
Basler M, Pilhofer M, Henderson GP, et al. (2012) Type VI secretion requires a dynamic contractile phage tail-like structure. Nature 483: 182-186. doi: 10.1038/nature10846
|
| [13] |
Kube S, Kapitein N, Zimniak T, et al. (2014) Structure of the VipA/B type VI secretion complex suggests a contraction-state-specific recycling mechanism. Cell Rep 8: 20-30. doi: 10.1016/j.celrep.2014.05.034
|
| [14] |
Clemens DL, Ge P, Lee BY, et al. (2015) Atomic structure of T6SS reveals interlaced array essential to function. Cell 160: 940-951. doi: 10.1016/j.cell.2015.02.005
|
| [15] |
Kudryashev M, Wang RY, Brackmann M, et al. (2015) Structure of the type VI secretion system contractile sheath. Cell 160: 952-962. doi: 10.1016/j.cell.2015.01.037
|
| [16] |
Heymann JB, Bartho JD, Rybakova D, et al. (2013) Three-dimensional structure of the toxin-delivery particle antifeeding prophage of Serratia entomophila. J Biol Chem 288: 25276-25284. doi: 10.1074/jbc.M113.456145
|
| [17] |
Shikuma NJ, Pilhofer M, Weiss GL, et al. (2014) Marine tubeworm metamorphosis induced by arrays of bacterial phage tail-like structures. Science 343: 529-533. doi: 10.1126/science.1246794
|
| [18] |
Veesler D, Cambillau C (2011) A common evolutionary origin for tailed-bacteriophage functional modules and bacterial machineries. Microbiol Mol Biol Rev 75: 423-433. doi: 10.1128/MMBR.00014-11
|
| [19] |
Leiman PG, Arisaka F, van Raaij MJ, et al. (2010) Morphogenesis of the T4 tail and tail fibers. Virology 7: 355. doi: 10.1186/1743-422X-7-355
|
| [20] |
Silverman JM, Brunet YR, Cascales E, et al. (2012) Structure and regulation of the type VI secretion system. Annu Rev Microbiol 66: 453-472. doi: 10.1146/annurev-micro-121809-151619
|
| [21] |
Ho BT, Dong TG, Mekalanos JJ (2014) A view to a kill: the bacterial type VI secretion system. Cell Host Microbe 15: 9-21. doi: 10.1016/j.chom.2013.11.008
|
| [22] |
Zoued A, Brunet YR, Durand E, et al. (2014) Architecture and assembly of the Type VI secretion system. Biochim Biophys Acta Mol Cell Res 1843: 1664-1673. doi: 10.1016/j.bbamcr.2014.03.018
|
| [23] |
Nakayama K, Takashima K, Ishihara H, et al. (2000) The R-type pyocin of Pseudomonas aeruginosa is related to P2 phage, and the F-type is related to lambda phage. Mol Microbiol 38: 213-231. doi: 10.1046/j.1365-2958.2000.02135.x
|
| [24] |
Liu Y, Schmidt B, Maskell DL (2010) MSAProbs: multiple sequence alignment based on pair hidden Markov models and partition function posterior probabilities. Bioinformatics 26: 1958-1964. doi: 10.1093/bioinformatics/btq338
|
| [25] | Felsenstein J (1989) Phylip: phylogeny inference package (version 3.2). Cladistics 5: 164-166. |
| [26] | Ackermann HW (2006) Classification of bacteriophages. In: Calendar R, editor. The Bacteriophages. New York, USA: Oxford University Press. 8-16. |
| [27] | Orlova EV (2012) Bacteriophages and their structural organisation. Bacteriophages 3-30. |
| [28] |
Ruska H (1942) Morphologische Befunde bei der bakteriophagen Lyse. Arch Gesamte Virusforsch 2: 345-387. doi: 10.1007/BF01249917
|
| [29] |
Ackermann HW (2011) The first phage electron micrographs. Bacteriophage 1: 225-227. doi: 10.4161/bact.1.4.17280
|
| [30] |
De Rosier DJ, Klug A (1968) Reconstruction of three dimensional structures from electron micrographs. Nature 217: 130-134. doi: 10.1038/217130a0
|
| [31] | Jacob F (1954) Induced biosynthesis and mode of action of a pyocine, antibiotic produced by Pseudomonas aeruginosa. Annales de l'Institut Pasteur 86: 149-160. |
| [32] |
Takeya K, Mlnamishima Y, Amako K, et al. (1967) A small rod-shaped pyocin. Virology 31: 166-168. doi: 10.1016/0042-6822(67)90021-9
|
| [33] |
Ishii SI, Nishi Y, Egami F (1965) The fine structure of a pyocin. J Mol Biol 13: 428-431. doi: 10.1016/S0022-2836(65)80107-3
|
| [34] | Shinomiya T, Shiga S, Kageyama M (1983) Genetic determinant of pyocin R2 in Pseudomonas aeruginosa PAO. I. Localization of the pyocin R2 gene cluster between the trpCD and trpE genes. Mol Gen Genet 189: 375-381. |
| [35] | Birmingham VA, Pattee PA (1981) Genetic transformation in Staphylococcus aureus: isolation and characterization of a competence-conferring factor from bacteriophage 80 alpha lysates. J Bacteriol 148: 301-307. |
| [36] |
Coetzee HL, de Klerk HC, Coetzee JN, et al. (1968) Bacteriophage-tail-like Particles Associated with Intra-species Killing of Proteus vulgaris. J Gen Virol 2: 29-36. doi: 10.1099/0022-1317-2-1-29
|
| [37] |
Gebhart D, Williams SR, Bishop-Lilly KA, et al. (2012) Novel high-molecular-weight, R-type bacteriocins of Clostridium difficile. J Bacteriol 194: 6240-6247. doi: 10.1128/JB.01272-12
|
| [38] | Matsui H, Sano Y, Ishihara H, et al. (1993) Regulation of pyocin genes in Pseudomonas aeruginosa by positive (prtN) and negative (prtR) regulatory genes. J Bacteriol 175: 1257-1263. |
| [39] |
Scholl D, Cooley M, Williams SR, et al. (2009) An engineered R-type pyocin is a highly specific and sensitive bactericidal agent for the food-borne pathogen Escherichia coli O157:H7. Antimicrob Agents Chemother 53: 3074-3080. doi: 10.1128/AAC.01660-08
|
| [40] |
Williams SR, Gebhart D, Martin DW, et al. (2008) Retargeting R-type pyocins to generate novel bactericidal protein complexes. Appl Environ Microbiol 74: 3868-3876. doi: 10.1128/AEM.00141-08
|
| [41] | Uratani Y, Hoshino T (1984) Pyocin R1 inhibits active transport inPseudomonas aeruginosa and depolarizes membrane potential. J Bacteriol 157: 632-636. |
| [42] |
Strauch E, Kaspar H, Schaudinn C, et al. (2001) Characterization of Enterocoliticin, a Phage Tail-Like Bacteriocin, and Its Effect on Pathogenic Yersinia enterocolitica Strains. Appl Environ Microbiol 67: 5634-5642. doi: 10.1128/AEM.67.12.5634-5642.2001
|
| [43] |
Ito S, Kageyama M, Egami F (1970) Isolation and characterization of pyocins from several strains of Pseudomonas aeruginosa. J Gen Appl Microbiol 16: 205-214. doi: 10.2323/jgam.16.3_205
|
| [44] |
Rodou A, Ankrah DO, Stathopoulos C (2010) Toxins and secretion systems of Photorhabdus luminescens. Toxins 2: 1250-1264. doi: 10.3390/toxins2061250
|
| [45] |
Yang G, Dowling AJ, Gerike U, et al. (2006) Photorhabdus virulence cassettes confer injectable insecticidal activity against the wax moth. J Bacteriol 188: 2254-2261. doi: 10.1128/JB.188.6.2254-2261.2006
|
| [46] |
Hurst MRH, Beard SS, Jackson TA, et al. (2007) Isolation and characterization of the Serratia entomophila antifeeding prophage. FEMS Microbiol Lett 270: 42-48. doi: 10.1111/j.1574-6968.2007.00645.x
|
| [47] |
Rybakova D, Radjainia M, Turner A, et al. (2013) Role of antifeeding prophage (Afp) protein Afp16 in terminating the length of the Afp tailocin and stabilizing its sheath. Mol Microbiol 89: 702-714. doi: 10.1111/mmi.12305
|
| [48] | Rybakova D, Schramm P, Mitra AK, et al. (2015) Afp14 is involved in regulating the length of Anti-feeding prophage (Afp). Mol Microbiol. |
| [49] | Ogata S, Suenaga H, Hayashida S (1982) Pock Formation of Streptomycetes endus with Production of Phage Taillike Particles. Appl Environ Microbiol 43: 1182-1187. |
| [50] |
Mougous JD, Cuff ME, Raunser S, et al. (2006) A virulence locus of Pseudomonas aeruginosa encodes a protein secretion apparatus. Science 312: 1526-1530. doi: 10.1126/science.1128393
|
| [51] |
Pukatzki S, Ma AT, Sturtevant D, et al. (2006) Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc Natl Acad Sci USA 103: 1528-1533. doi: 10.1073/pnas.0510322103
|
| [52] |
Bingle LE, Bailey CM, Pallen MJ (2008) Type VI secretion: a beginner's guide. Curr Opin Microbiol 11: 3-8. doi: 10.1016/j.mib.2008.01.006
|
| [53] |
Boyer F, Fichant G, Berthod J, et al. (2009) Dissecting the bacterial type VI secretion system by a genome wide in silico analysis: what can be learned from available microbial genomic resources? BMC Genomics 10: 104. doi: 10.1186/1471-2164-10-104
|
| [54] | Bröms JE, Sjöstedt A, Lavander M (2010) The Role of the Francisella tularensis Pathogenicity Island in Type VI Secretion, Intracellular Survival, and Modulation of Host Cell Signaling. Front Microbiol 1: 136-136. |
| [55] | de Bruin OM, Duplantis BN, Ludu JS, et al. (2011) The biochemical properties of the Francisella Pathogenicity Island (FPI)-encoded proteins, IglA, IglB, IglC, PdpB and DotU, suggest roles in type VI secretion. Microbiology. |
| [56] |
Russell AB, Wexler AG, Harding BN, et al. (2014) A type VI secretion-related pathway in Bacteroidetes mediates interbacterial antagonism. Cell Host Microbe 16: 227-236. doi: 10.1016/j.chom.2014.07.007
|
| [57] |
Shalom G, Shaw JG, Thomas MS (2007) In vivo expression technology identifies a type VI secretion system locus in Burkholderia pseudomallei that is induced upon invasion of macrophages. Microbiology 153: 2689-2699. doi: 10.1099/mic.0.2007/006585-0
|
| [58] |
Leiman PG, Basler M, Ramagopal UA, et al. (2009) Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc Natl Acad Sci USA 106: 4154-4159. doi: 10.1073/pnas.0813360106
|
| [59] |
Pell LG, Kanelis V, Donaldson LW, et al. (2009) The phage lambda major tail protein structure reveals a common evolution for long-tailed phages and the type VI bacterial secretion system. Proc Natl Acad Sci USA 106: 4160-4165. doi: 10.1073/pnas.0900044106
|
| [60] |
Lossi NS, Dajani R, Freemont P, et al. (2011) Structure-function analysis of HsiF, a gp25-like component of the type VI secretion system, in Pseudomonas aeruginosa. Microbiology 157: 3292-3305. doi: 10.1099/mic.0.051987-0
|
| [61] |
Bonemann G, Pietrosiuk A, Diemand A, et al. (2009) Remodelling of VipA/VipB tubules by ClpV-mediated threading is crucial for type VI protein secretion. EMBO J 28: 315-325. doi: 10.1038/emboj.2008.269
|
| [62] |
Lossi NS, Manoli E, Forster A, et al. (2013) The HsiB1C1 (TssB-TssC) complex of the Pseudomonas aeruginosa type VI secretion system forms a bacteriophage tail sheathlike structure. J Biol Chem 288: 7536-7548. doi: 10.1074/jbc.M112.439273
|
| [63] |
Zheng J, Ho B, Mekalanos JJ (2011) Genetic analysis of anti-amoebae and anti-bacterial activities of the type VI secretion system in Vibrio cholerae. PLoS One 6: e23876-e23876. doi: 10.1371/journal.pone.0023876
|
| [64] |
Zoued A, Durand E, Bebeacua C, et al. (2013) TssK is a trimeric cytoplasmic protein interacting with components of both phage-like and membrane anchoring complexes of the type VI secretion system. J Biol Chem 288: 27031-27041. doi: 10.1074/jbc.M113.499772
|
| [65] |
Durand E, Zoued A, Spinelli S, et al. (2012) Structural characterization and oligomerization of the TssL protein, a component shared by bacterial type VI and type IVb secretion systems. J Biol Chem 287: 14157-14168. doi: 10.1074/jbc.M111.338731
|
| [66] |
Robb CS, Nano FE, Boraston AB (2012) The structure of the conserved type six secretion protein TssL (DotU) from Francisella novicida. J Mol Biol 419: 277-283. doi: 10.1016/j.jmb.2012.04.003
|
| [67] |
Ma LS, Lin JS, Lai EM (2009) An IcmF family protein, ImpLM, is an integral inner membrane protein interacting with ImpKL, and its walker a motif is required for type VI secretion system-mediated Hcp secretion in Agrobacterium tumefaciens. J Bacteriol 191: 4316-4329. doi: 10.1128/JB.00029-09
|
| [68] | Das S, Chaudhuri K (2003) Identification of a unique IAHP (IcmF associated homologous proteins) cluster in Vibrio cholerae and other proteobacteria through in silico analysis. In Silico Biol 3: 287-300. |
| [69] |
Felisberto-Rodrigues C, Durand E, Aschtgen MS, et al. (2011) Towards a Structural Comprehension of Bacterial Type VI Secretion Systems: Characterization of the TssJ-TssM Complex of an Escherichia coli Pathovar. PLoS Pathog 7: e1002386. doi: 10.1371/journal.ppat.1002386
|
| [70] |
Rao VA, Shepherd SM, English G, et al. (2011) The structure of Serratia marcescens Lip, a membrane-bound component of the type VI secretion system. Acta Crystallogr D Biol Crystallogr 67: 1065-1072. doi: 10.1107/S0907444911046300
|
| [71] |
Miyata ST, Bachmann V, Pukatzki S (2013) Type VI secretion system regulation as a consequence of evolutionary pressure. J Med Microbiol 62: 663-676. doi: 10.1099/jmm.0.053983-0
|
| [72] |
Bernard CS, Brunet YR, Gavioli M, et al. (2011) Regulation of type VI secretion gene clusters by sigma54 and cognate enhancer binding proteins and cognate enhancer binding proteins. Journal of bacteriology 193: 2158-2167. doi: 10.1128/JB.00029-11
|
| [73] |
Kitaoka M, Miyata ST, Brooks TM, et al. (2011) VasH is a transcriptional regulator of the type VI secretion system functional in endemic and pandemicVibrio cholerae. J Bacteriology 193: 6471-6482. doi: 10.1128/JB.05414-11
|
| [74] |
Dong TG, Mekalanos JJ (2012) Characterization of the RpoN regulon reveals differential regulation of T6SS and new flagellar operons in Vibrio cholerae O37 strain V52. Nucleic acids research 40: 7766-7775. doi: 10.1093/nar/gks567
|
| [75] |
Basler M, Ho BT, Mekalanos JJ (2013) Tit-for-tat: type VI secretion system counterattack during bacterial cell-cell interactions. Cell 152: 884-894. doi: 10.1016/j.cell.2013.01.042
|
| [76] |
Basler M, Mekalanos JJ (2012) Type 6 secretion dynamics within and between bacterial cells. Science 337: 815. doi: 10.1126/science.1222901
|
| [77] |
Ho BT, Basler M, Mekalanos JJ (2013) Type 6 secretion system-mediated immunity to type 4 secretion system-mediated gene transfer. Science 342: 250-253. doi: 10.1126/science.1243745
|
| [78] |
Fritsch MJ, Trunk K, Diniz JA, et al. (2013) Proteomic Identification of Novel Secreted Antibacterial Toxins of the Serratia marcescens Type VI Secretion System. Mol Cell Proteomics 12: 2735-2749. doi: 10.1074/mcp.M113.030502
|
| [79] |
Mougous JD, Gifford CA, Ramsdell TL, et al. (2007) Threonine phosphorylation post-translationally regulates protein secretion in Pseudomonas aeruginosa. Nat Cell Biol 9: 797-803. doi: 10.1038/ncb1605
|
| [80] |
Kapitein N, Bonemann G, Pietrosiuk A, et al. (2013) ClpV recycles VipA/VipB tubules and prevents non-productive tubule formation to ensure efficient type VI protein secretion. Mol Microbiol 87: 1013-1028. doi: 10.1111/mmi.12147
|
| [81] |
Hsu F, Schwarz S, Mougous JD (2009) TagR promotes PpkA-catalysed type VI secretion activation in Pseudomonas aeruginosa. Mol Microbiol 72: 1111-1125. doi: 10.1111/j.1365-2958.2009.06701.x
|
| [82] |
Silverman JM, Austin LS, Hsu F, et al. (2011) Separate inputs modulate phosphorylation-dependent and -independent type VI secretion activation. Mol Microbiol 82: 1277-1290. doi: 10.1111/j.1365-2958.2011.07889.x
|
| [83] |
Casabona MG, Silverman JM, Sall KM, et al. (2013) An ABC transporter and an outer membrane lipoprotein participate in posttranslational activation of type VI secretion in Pseudomonas aeruginosa. Environ Microbiol 15: 471-486. doi: 10.1111/j.1462-2920.2012.02816.x
|
| [84] |
Lin JS, Wu HH, Hsu PH, et al. (2014) Fha interaction with phosphothreonine of TssL activates type VI secretion in Agrobacterium tumefaciens. PLoS Pathog 10: e1003991-e1003991. doi: 10.1371/journal.ppat.1003991
|
| [85] |
Zheng J, Leung KY (2007) Dissection of a type VI secretion system in Edwardsiella tarda. Mol Microbiol 66: 1192-1206. doi: 10.1111/j.1365-2958.2007.05993.x
|
| [86] |
Ma LS, Narberhaus F, Lai EM (2012) IcmF family protein TssM exhibits ATPase activity and energizes type VI secretion. J Biol Chem 287: 15610-15621. doi: 10.1074/jbc.M111.301630
|
| [87] |
Shneider MM, Buth SA, Ho BT, et al. (2013) PAAR-repeat proteins sharpen and diversify the type VI secretion system spike. Nature 500: 350-353. doi: 10.1038/nature12453
|
| [88] |
Brunet YR, Hénin J, Celia H, et al. (2014) Type VI secretion and bacteriophage tail tubes share a common assembly pathway. EMBO Rep 15: 315-321. doi: 10.1002/embr.201337936
|
| [89] |
Bröms JE, Lavander M, Sjöstedt A (2009) A conserved alpha-helix essential for a type VI secretion-like system of Francisella tularensis. J Bacteriol 191: 2431-2446. doi: 10.1128/JB.01759-08
|
| [90] |
Aubert DF, MacDonald DK, Valvano MA (2010) BcsKC is an essential protein for the type VI secretion system activity in Burkholderia cenocepacia that forms an outer membrane complex with BcsLB. J Biol Chem 285: 35988-35998. doi: 10.1074/jbc.M110.120402
|
| [91] |
Chang YW, Chen S, Tocheva EI, et al. (2014) Correlated cryogenic photoactivated localization microscopy and cryo-electron tomography. Nature Methods 11: 737-739. doi: 10.1038/nmeth.2961
|
| [92] |
King J (1971) Bacteriophage T4 tail assembly: four steps in core formation. J Mol Biol 58: 693-709. doi: 10.1016/0022-2836(71)90034-9
|
| [93] |
Silverman JM, Agnello DM, Zheng H, et al. (2013) Haemolysin coregulated protein is an exported receptor and chaperone of type VI secretion substrates. Mol Cell 51: 584-593. doi: 10.1016/j.molcel.2013.07.025
|
| [94] |
Brooks TM, Unterweger D, Bachmann V, et al. (2013) Lytic activity of the Vibrio cholerae type VI secretion toxin VgrG-3 is inhibited by the antitoxin TsaB. J Biol Chem 288: 7618-7625. doi: 10.1074/jbc.M112.436725
|
| [95] |
Pietrosiuk A, Lenherr ED, Falk S, et al. (2011) Molecular basis for the unique role of the AAA+ chaperone ClpV in type VI protein secretion. J Biol Chem 286: 30010-30021. doi: 10.1074/jbc.M111.253377
|
| [96] |
Forster A, Planamente S, Manoli E, et al. (2014) Coevolution of the ATPase ClpV, the Sheath Proteins TssB and TssC and the Accessory Protein TagJ/HsiE1 Distinguishes Type VI Secretion Classes. J Biol Chem 289: 33032-33043. doi: 10.1074/jbc.M114.600510
|
| [97] |
King J (1968) Assembly of the tail of bacteriophage T4. J Mol Biol 32: 231-262. doi: 10.1016/0022-2836(68)90007-7
|
| [98] |
Abuladze NK, Gingery M, Tsai J, et al. (1994) Tail length determination in bacteriophage T4. Virology 199: 301-310. doi: 10.1006/viro.1994.1128
|
| [99] |
Kanamaru S, Leiman PG, Kostyuchenko VA, et al. (2002) Structure of the cell-puncturing device of bacteriophage T4. Nature 415: 553-557. doi: 10.1038/415553a
|
| [100] |
Kanamaru S, Ishiwata Y, Suzuki T, et al. (2005) Control of bacteriophage T4 tail lysozyme activity during the infection process. J Mol Biol 346: 1013-1020. doi: 10.1016/j.jmb.2004.12.042
|
| [101] |
Pukatzki S, Ma AT, Revel AT, et al. (2007) Type VI secretion system translocates a phage tail spike-like protein into target cells where it cross-links actin. Proc Natl Acad Sci USA 104: 15508-15513. doi: 10.1073/pnas.0706532104
|
| [102] |
Ma AT, McAuley S, Pukatzki S, et al. (2009) Translocation of a Vibrio cholerae type VI secretion effector requires bacterial endocytosis by host cells. Cell Host Microbe 5: 234-243. doi: 10.1016/j.chom.2009.02.005
|
| [103] |
Suarez G, Sierra JC, Erova TE, et al. (2010) A type VI secretion system effector protein, VgrG1, from Aeromonas hydrophila that induces host cell toxicity by ADP ribosylation of actin. J Bacteriol 192: 155-168. doi: 10.1128/JB.01260-09
|
| [104] |
Durand E, Derrez E, Audoly G, et al. (2012) Crystal structure of the VgrG1 actin cross-linking domain of the Vibrio cholerae type VI secretion system. J Biol Chem 287: 38190-38199. doi: 10.1074/jbc.M112.390153
|
| [105] |
Browning C, Shneider MM, Bowman VD, et al. (2012) Phage pierces the host cell membrane with the iron-loaded spike. Structure 20: 326-339. doi: 10.1016/j.str.2011.12.009
|
| [106] |
Moody MF, Makowski L (1981) X-ray diffraction study of tail-tubes from bacteriophage T2L. J Mol Biol 150: 217-244. doi: 10.1016/0022-2836(81)90450-2
|
| [107] |
Wagenknecht T, Bloomfield VA (1977) In vitro polymerization of bacteriophage T4D tail core subunits. J Mol Biol 116: 347-359. doi: 10.1016/0022-2836(77)90074-2
|
| [108] |
Poglazov BF, Nikolskaya TI (1969) Self-assembly of the protein of bacteriophage T2 tail cores. J Mol Biol 43: 231-233. doi: 10.1016/0022-2836(69)90094-1
|
| [109] |
Douzi B, Spinelli S, Blangy S, et al. (2014) Crystal structure and self-interaction of the type VI secretion tail-tube protein from enteroaggregative Escherichia coli. PLoS One 9: e86918-e86918. doi: 10.1371/journal.pone.0086918
|
| [110] |
Lin JS, Ma LS, Lai EM (2013) Systematic dissection of the agrobacterium type VI secretion system reveals machinery and secreted components for subcomplex formation. PLoS One 8: e67647-e67647. doi: 10.1371/journal.pone.0067647
|
| [111] |
Kostyuchenko VA, Leiman PG, Chipman PR, et al. (2003) Three-dimensional structure of bacteriophage T4 baseplate. Nat Struct Mol Biol 10: 688-693. doi: 10.1038/nsb970
|
| [112] |
Lim YT, Jobichen C, Wong J, et al. (2015) Extended loop region of Hcp1 is critical for the assembly and function of type VI secretion system in Burkholderia pseudomallei. Sci Rep 5: 8235. doi: 10.1038/srep08235
|
| [113] |
Jobichen C, Chakraborty S, Li M, et al. (2010) Structural basis for the secretion of EvpC: a key type VI secretion system protein from Edwardsiella tarda. PLoS One 5: e12910. doi: 10.1371/journal.pone.0012910
|
| [114] |
Aksyuk AA, Leiman PG, Kurochkina LP, et al. (2009) The tail sheath structure of bacteriophage T4: a molecular machine for infecting bacteria. EMBO J 28: 821-829. doi: 10.1038/emboj.2009.36
|
| [115] |
Fokine A, Zhang Z, Kanamaru S, et al. (2013) The molecular architecture of the bacteriophage T4 neck. J Mol Biol 425: 1731-1744. doi: 10.1016/j.jmb.2013.02.012
|
| [116] |
Broms JE, Ishikawa T, Wai SN, et al. (2013) A functional VipA-VipB interaction is required for the type VI secretion system activity of Vibrio cholerae O1 strain A1552. BMC Microbiol 13: 96. doi: 10.1186/1471-2180-13-96
|
| [117] |
Zhang XY, Brunet YR, Logger L, et al. (2013) Dissection of the TssB-TssC Interface during Type VI Secretion Sheath Complex Formation. PLoS One 8: e81074. doi: 10.1371/journal.pone.0081074
|
| [118] |
Poglazov BF, Efimov AV, Marco S, et al. (1999) Polymerization of bacteriophage T4 tail sheath protein mutants truncated at the C-termini. J Struct Biol 127: 224-230. doi: 10.1006/jsbi.1999.4164
|
| [119] | Moody MF (1973) Sheath of bacteriophage T4. 3. Contraction mechanism deduced from partially contracted sheaths. J Mol Biol 80: 613-635. |
| [120] |
Efimov AV, Kurochkina LP, Mesyanzhinov VV (2002) Engineering of bacteriophage T4 tail sheath protein. Biochemistry (Moscow) 67: 1366-1370. doi: 10.1023/A:1021857926152
|
| [121] |
Takeda S, Suzuki M, Yamada T, et al. (2004) Mapping of functional sites on the primary structure of the contractile tail sheath protein of bacteriophage T4 by mutation analysis. Biochim Biophys Acta, Proteins Proteom 1699: 163-171. doi: 10.1016/S1570-9639(04)00058-5
|
| [122] |
Maxwell KL, Fatehi HM, Chang T, et al. (2013) Structural and functional studies of gpX of Escherichia coli phage P2 reveal a widespread role for LysM domains in the baseplates of contractile-tailed phages. J Bacteriol 195: 5461-5468. doi: 10.1128/JB.00805-13
|
| [123] |
Bradley DE (1963) The structure of coliphages. J Gen Microbiol 31: 435-445. doi: 10.1099/00221287-31-3-435
|
| [124] |
Leblanc C, Caumont-Sarcos A, Comeau AM, et al. (2009) Isolation and genomic characterization of the first phage infecting Iodobacteria: varphiPLPE, a myovirus having a novel set of features. Env Microbiol Rep 1: 499-509. doi: 10.1111/j.1758-2229.2009.00055.x
|
| [125] | Moody MF (1967) Structure of the sheath of bacteriophage T4. I. Structure of the contracted sheath and polysheath. J Mol Biol 25: 167-200. |
| [126] | Moody MF (1967) Structure of the sheath of bacteriophage T4. II. Rearrangement of the sheath subunits during contraction. J Mol Biol 25: 201-208. |
| [127] |
Uratani Y (1982) A circular dichroism study of sheath contraction in pyocin R1. Biochim Biophys Acta 703: 196-203. doi: 10.1016/0167-4838(82)90048-6
|
| [128] |
Venyaminov SY, Rodikova LP, Metlina AL, et al. (1975) Secondary structure change of bacteriophage T4 sheath protein during sheath contraction. J Mol Biol 98: 657-664. doi: 10.1016/S0022-2836(75)80001-5
|
| [129] |
Bai X-c, McMullan G, Scheres SHW (2015) How cryo-EM is revolutionizing structural biology. Trends Biochem Sci 40: 49-57. doi: 10.1016/j.tibs.2014.10.005
|
| [130] |
Pettersen EF, Goddard TD, Huang CC, et al. (2004) UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 25: 1605-1612. doi: 10.1002/jcc.20084
|
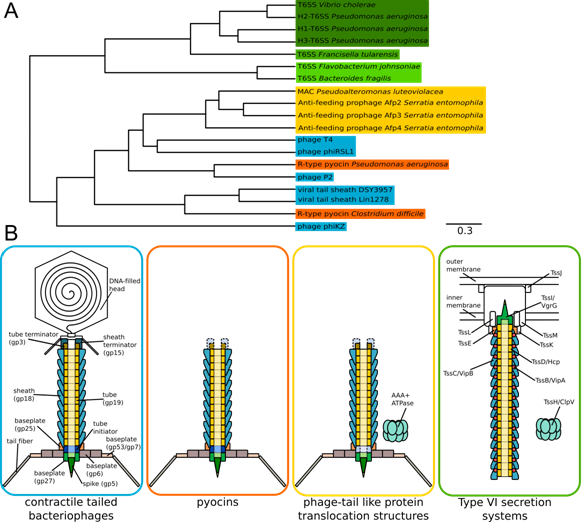
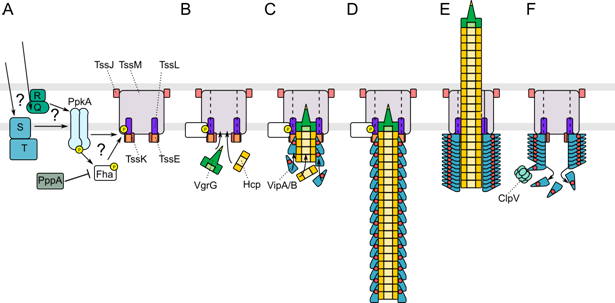
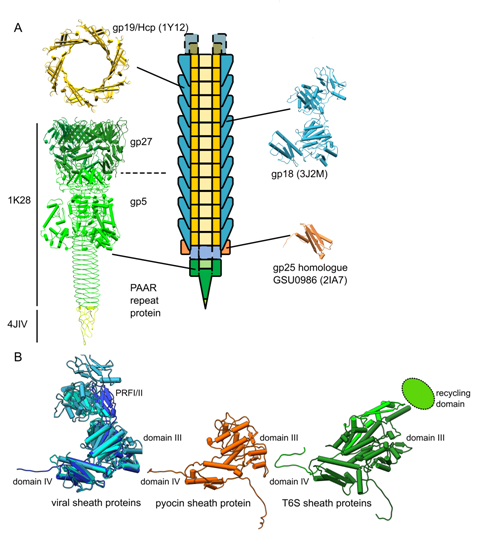
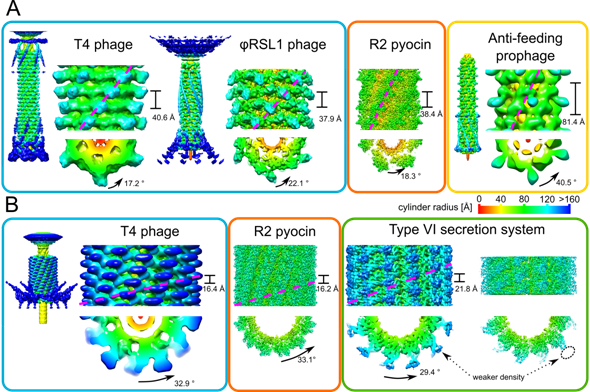

Figures(7) / Tables(2)

Sebastian Kube, Petra Wendler. Structural comparison of contractile nanomachines[J]. AIMS Biophysics, 2015, 2(2): 88-115. doi: 10.3934/biophy.2015.2.88







 DownLoad:
DownLoad: