Citation: Thatiane Rodrigues Mota, Dyoni Matias de Oliveira, Rogério Marchiosi, Osvaldo Ferrarese-Filho, Wanderley Dantas dos Santos. Plant cell wall composition and enzymatic deconstruction[J]. AIMS Bioengineering, 2018, 5(1): 63-77. doi: 10.3934/bioeng.2018.1.63

| [1] | Santos WDD, Gómez ED, Buckeridge MS, (2011) Bioenergy and the Sustainable Revolution, In: Buckeridge MS, Goldman GH. Routes to Cellulosic Ethanol. Springer Science and Business Media, LCC, 1 Ed., New York, 15–26. |
| [2] |
Jaiswal D, Souza APD, Larsen S, et al. (2017) Brazilian sugarcane ethanol as an expandable green alternative to crude oil use. Nat Clim Change 7: 788–792. doi: 10.1038/nclimate3410

|
| [3] | Saini JK, Saini R, Tewari L (2015) Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: Concepts and recent developments. Biotech 5: 337–353. |
| [4] | Buckeridge MS, Dos Santos WD, Mas T, et al. (2016) The cell wall architecture of sugarcane and its implications to cell wall recalcitrance, In: Lam E, Carrer H, Da Silva JA, et al. Compendium of Bioenergy Plants: Compendium of Bioenergy Plants, Boca Raton, FL: CRC Press, 31–50. |
| [5] |
Poovaiah CR, Nageswara-Rao M, Soneji JR, et al. (2014) Altered lignin biosynthesis using biotechnology to improve lignocellulosic biofuel feedstocks. Plant Biotechnol J 12: 1163–1173. doi: 10.1111/pbi.12225
|
| [6] |
Torres AF, Visser RGF, Trindade LM (2015) Bioethanol from maize cell walls: Genes, molecular tools, and breeding prospects. GCB Bioenergy 7: 591–607. doi: 10.1111/gcbb.12164
|
| [7] |
Balat M (2011) Production of bioethanol from lignocellulosic materials via the biochemical pathway: A review. Energ Convers Manage 52: 858–875. doi: 10.1016/j.enconman.2010.08.013
|
| [8] |
Souza APD, Leite DCC, Pattathil S, et al. (2013) Composition and structure of sugarcane cell wall polysaccharides: Implications for second-generation bioethanol production. BioEnergy Res 6: 564–579. doi: 10.1007/s12155-012-9268-1
|
| [9] |
Gupta A, Verma JP (2015) Sustainable bio-ethanol production from agro-residues. Renew Sust Energ Rev 41: 550–567. doi: 10.1016/j.rser.2014.08.032
|
| [10] |
Sims REH, Mabee W, Saddler JN, et al. (2010) An overview of second generation biofuel technologies. Bioresource Technol 101: 1570–1580. doi: 10.1016/j.biortech.2009.11.046
|
| [11] |
Amorim HV, Lopes ML, Oliveira JVC, et al. (2011) Scientific challenges of bioethanol production in Brazil. Appl Microbiol Biotechnol 91: 1267–1275. doi: 10.1007/s00253-011-3437-6
|
| [12] | Verardi A, Blasi A, Molino A, et al. (2016) Improving the enzymatic hydrolysis of Saccharum officinarum L. bagasse by optimizing mixing in a stirred tank reactor: Quantitative analysis of biomass conversion. Fuel Process Technol 149: 15–22. |
| [13] |
Limayem A, Ricke SC (2012) Lignocellulosic biomass for bioethanol production: Current perspectives, potential issues and future prospects. Prog Energy Combust Sci 38: 449–467. doi: 10.1016/j.pecs.2012.03.002
|
| [14] |
Goldemberg J, Coelho ST, Guardabassi P (2008) The sustainability of ethanol production from sugarcane. Energ Policy 36: 2086–2097. doi: 10.1016/j.enpol.2008.02.028
|
| [15] | Rocha GJM, Gonçalves AR, Oliveira BR, et al. (2013) Steam explosion pretreatment reproduction and alkaline delignification reactions performed on a pilot scale with sugarcane bagasse for bioethanol production. Ind Crop Prod 35: 274–279. |
| [16] | Buckeridge MS, Santos WDD, Souza AP, (2010) Routes for cellulosic ethanol in Brazil, In: Cortez LAB. Sugarcane Bioethanol, R&D for productivity and sustainability, Blucher, 992. |
| [17] | Kou L, Song Y, Zhang X, et al. (2017) Comparison of four types of energy grasses as lignocellulosic feedstock for the production of bio-ethanol. Bioresource Technol 241:434–429. |
| [18] |
Álvarez C, Manuel RF, Bruno D (2016) Enzymatic hydrolysis of biomass from wood. Microb Biotechnol 9: 149–156. doi: 10.1111/1751-7915.12346
|
| [19] | Malinovsky FG, Fangel JU, Willats WGT (2014) The role of the cell wall in plant immunity. Front Plant Sci 5: 178. |
| [20] |
Payne CM, Knott BC, Mayes HB, et al. (2015) Fungal cellulases. Chem Rev 115: 1308–1448. doi: 10.1021/cr500351c
|
| [21] |
Khare SK, Pandey A, Larroche C (2015) Current perspectives in enzymatic saccharification of lignocellulosic biomass. Biochem Eng J 102: 38–44. doi: 10.1016/j.bej.2015.02.033
|
| [22] | Jørgensen H, Kristensen JB, Felby C (2007) Enzymatic conversion of lignocellulose into fermentable sugars: Challenges and opportunities. Biofuels Bioprod Biorefin 1: 119–134. |
| [23] |
Le GH, Florian F, Jean-Marc D, et al. (2015) Cell wall metabolism in response to abiotic stress. Plants 4: 112–166. doi: 10.3390/plants4010112
|
| [24] |
Pauly M, Keegstra K (2008) Cell-wall carbohydrates and their modification as are source for biofuels. Plant J 54: 559–568. doi: 10.1111/j.1365-313X.2008.03463.x
|
| [25] |
Höfte H, Voxeur A (2017) Plant cell walls. Curr Biol 27: R865–R870. doi: 10.1016/j.cub.2017.05.025
|
| [26] |
Carpita NC, Gibeaut DM (1993) Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth. Plant J 3: 1–30. doi: 10.1111/j.1365-313X.1993.tb00007.x
|
| [27] |
Chundawat SPS, Beckham GT, Himmel ME, et al. (2011) Deconstruction of lignocellulosic biomass to fuels and chemicals. Annu Rev Chem Biomol Eng 2: 121–145. doi: 10.1146/annurev-chembioeng-061010-114205
|
| [28] | Meents MJ, Watanabe Y, Samuels AL (2018) The cell biology of secondary cell wall biosynthesis. Ann Bot. |
| [29] | Turner S, Kumar M (2018) Cellulose synthase complex organization and cellulose microfibril structure. Philos Trans 376: 20170048. |
| [30] |
Himmel ME, Ding SY, Johnson DK, et al. (2007) Biomass recalcitrance: Engineering plants and enzymes for biofuels production. Science 315: 804–807. doi: 10.1126/science.1137016
|
| [31] | Carpita NC, Mccann MC, (2000) The cell wall, In: Buchanan BB, Gruissem W, Jones RL, Biochemistry and molecular biology of plants. Rockville: IL: American Society of Plant Physiologists, 1367. |
| [32] |
Abramson M, Shoseyov O, Shani Z (2010) Plant cell wall reconstruction toward improved lignocellulosic production and processability. Plant Sci 178: 61–72. doi: 10.1016/j.plantsci.2009.11.003
|
| [33] | Van dWT, Dolstra O, Visser RG, et al. (2017) Stability of cell wall composition and saccharification efficiency in miscanthus across diverse environments. Front Plant Sci 7: 2004. |
| [34] |
Pauly M, Keegstra K (2016) Biosynthesis of the plant cell wall matrix polysaccharide xyloglucan. Annu Rev Plant Biol 67: 235–259. doi: 10.1146/annurev-arplant-043015-112222
|
| [35] | Oliveira DM, Finger-Teixeira A, Mota TR, et al. (2016). Ferulic acid: A key component in grass lignocellulose recalcitrance to hydrolysis. Plant Biotechnol J 13: 1224–1232. |
| [36] |
Kozlova LV, Ageeva MV, Ibragimova NN, et al. (2014) Arrangement of mixed-linkage glucan and glucuronoarabinoxylan in the cell walls of growing maize roots. Ann Bot 114: 1135–1145. doi: 10.1093/aob/mcu125
|
| [37] |
Srivastava PK, Kapoor M (2017) Production, properties, and applications of endo-β-mannanases. Biotechnol Adv 35: 1–19. doi: 10.1016/j.biotechadv.2016.11.001
|
| [38] | Biswal AK, Atmodjo MA, Li M, et al. (2018) Sugar release and growth of biofuel crops are improved by downregulation of pectin biosynthesis. Nat Biotechnol. |
| [39] |
Voragen AGJ, Coenen GJ, Verhoef RP, et al. (2009) Pectin, a versatile polysaccharide present in plant cell walls. Struct Chem 20: 263–275. doi: 10.1007/s11224-009-9442-z
|
| [40] |
Mohnen D (2008) Pectin structure and biosynthesis. Curr Opin Plant Biol 11: 266–277. doi: 10.1016/j.pbi.2008.03.006
|
| [41] | Nguyen TN, Son S, Jordan MC, et al. (2016) Lignin biosynthesis in wheat (Triticum aestivum L.): Its response to waterlogging and association with hormonal levels. BMC Plant Biol 16: 1–16. |
| [42] |
Ragauskas AJ, Beckham GT, Biddy MJ, et al. (2014) Lignin valorization: Improving lignin processing in the biorefinery. Science 344: 1246843. doi: 10.1126/science.1246843
|
| [43] | Santos WDD, Ferrarese MDLL (2008) Ferulic acid: An allelochemical troublemaker. Funct Plant Sci Biotechnol 2: 47–55. |
| [44] |
dos Santos WD, Ferrarese ML, Nakamura CV, et al. (2008) Soybean (Glycine max) root lignification induced by ferulic acid the possible mode of action. J Chem Ecol 34: 1230–1241. doi: 10.1007/s10886-008-9522-3
|
| [45] | Liu Q, Luo L, Zheng L (2018) Lignins: Biosynthesis and Biological Functions in Plants. Int J Mol Sci 19: 335. |
| [46] |
Salvador VH, Lima RB, dos Santos WD, et al. (2013) Cinnamic acid increases lignin production and inhibits soybean root growth. PLoS One 8: e69105. doi: 10.1371/journal.pone.0069105
|
| [47] | Oliveira DMD, Finger-Teixeira A, Freitas DLD, et al. (2017) Phenolic Compounds in Plants: Implications for Bioenergy, In: Buckeridge MS, de Souza AP, Advances of Basic Science for Second Generation Bioethanol from Sugarcane, Springer Int Publishing, 39–52. |
| [48] | Boerjan W, Ralph J, Baucher M (2003) Lignin biosynthesis. Annu Rev Plant Biol 166: 63–71. |
| [49] |
Moreira-Vilar FC, Siqueira-Soares RC, Finger-Teixeira A, et al. (2014) The acetyl bromide method is faster, simpler and presents best recovery of lignin in different herbaceous tissues than klason and thioglycolic acid methods. PLoS One 9: e110000. doi: 10.1371/journal.pone.0110000
|
| [50] | Santos WDD, Marchiosi R, Vilar FCW, et al. (2014) Polyvalent Lignin: Recent Approaches in Determination and Applications, In: Lu F, Lignin: Structural analysis, applications in biomaterials and ecological significance, New York: Nova Publishers, 1–26. |
| [51] | Fornalé S, Rencoret J, Garcíacalvo L, et al. (2017) Changes in cell wall polymers and degradability in maize mutants lacking 3′- and 5′-O-methyltransferases involved in lignin biosynthesis. Plant Cell Physiol 58: 240–255. |
| [52] |
Lima RB, Salvador VH, dos Santos WD, et al. (2013) Enhanced lignin monomer production caused by cinnamic acid and its hydroxylated derivatives inhibits soybean root growth. PLoS One 8: e80542. doi: 10.1371/journal.pone.0080542
|
| [53] |
Saritha M, Arora A, Lata (2012) Biological Pretreatment of Lignocellulosic Substrates for Enhanced Delignification and Enzymatic Digestibility. Indian J Microbiol 52: 122–130. doi: 10.1007/s12088-011-0199-x
|
| [54] |
Henrissat B, Bairoch A (1993) New families in the classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem J 293: 781–788. doi: 10.1042/bj2930781
|
| [55] |
Lombard V, Ramulu HG, Drula E, et al. (2014) The carbohydrate-active enzymes database (CAZy). Nucleic Acids Res 42: D490–D495. doi: 10.1093/nar/gkt1178
|
| [56] |
Bourne Y, Henrissat B (2001) Glycoside hydrolases and glycosyltransferases families and functional modules. Curr Opin Struc Biol 11: 593–600. doi: 10.1016/S0959-440X(00)00253-0
|
| [57] |
Hashimoto H (2006) Recent structural studies of carbohydrate-binding modules. Cell Mol Life Sci 63: 2954–2967. doi: 10.1007/s00018-006-6195-3
|
| [58] |
Boraston AB, Bolam DN, Gilbert HJ, et al. (2004) Carbohydrate-binding modules: Fine-tuning polysaccharide recognition. Biochem J 382: 769–781. doi: 10.1042/BJ20040892
|
| [59] |
Davies G, Henrissat B (1995) Structures and mechanisms of glycosyl hydrolases. Structure 3: 853–859. doi: 10.1016/S0969-2126(01)00220-9
|
| [60] | Terrapon N, Lombard V, Drula E, et al. (2017) The CAZy Database/the Carbohydrate-Active Enzyme (CAZy) Database: Principles and Usage Guidelines, In: Aoki-Kinoshita K (eds), A Practical Guide to Using Glycomics Databases. Springer, Tokyo, 117–131. |
| [61] | Segato F, Dias B, Berto GL, et al. (2017) Cloning, heterologous expression and biochemical characterization of a non-specific endoglucanase family 12 from Aspergillus terreus NIH2624. BBA 1865: 395–403. |
| [62] |
Segato F, Damásio AR, de Lucas RC, et al. (2014) Genomics review of holocellulose deconstruction by aspergilli. Microbiol Mol Biol Rev 78: 588–613. doi: 10.1128/MMBR.00019-14
|
| [63] | Olsen JP, Alasepp K, Kari J, et al. (2016) Mechanism of product inhibition for cellobiohydrolase Cel7A during hydrolysis of insoluble cellulose. Biotechnol Bioeng 113: 1178–1186. |
| [64] |
Grange DC, Haan R, Zyl WH (2010) Engineering cellulolytic ability into bioprocessing organisms. Appl Microbiol Biotechnol 87: 1195–1208. doi: 10.1007/s00253-010-2660-x
|
| [65] |
Damásio ARL, Rubio MV, Gonçalves TA, et al. (2017) Xyloglucan breakdown by endo-xyloglucanase family 74 from Aspergillus fumigatus. Appl Microbiol Biotechnol 101: 2893–2903. doi: 10.1007/s00253-016-8014-6
|
| [66] |
Dodd D, Cann IKO (2009) Enzymatic deconstruction of xylan for biofuel production. GCB Bioenergy 1: 2–17. doi: 10.1111/j.1757-1707.2009.01004.x
|
| [67] |
Damásio ARDL, Pessedela BC, Segato F, et al. (2012) Improvement of fungal arabinofuranosidase thermal stability by reversible immobilization. Process Biochem 47: 2411–2417. doi: 10.1016/j.procbio.2012.09.024
|
| [68] |
Wilkens C, Andersen S, Dumon C, et al. (2017) GH62 arabinofuranosidases: Structure, function and applications. Biotechnol Adv 35: 792–804. doi: 10.1016/j.biotechadv.2017.06.005
|
| [69] |
Wu L, Jiang J, Kallemeijn W, et al. (2017) Activity-based probes for functional interrogation of retaining β-glucuronidases. Nat Chem Biol 13: 867–873 doi: 10.1038/nchembio.2395
|
| [70] |
Bornscheuer UT (2002) Microbial carboxyl esterases: Classification, properties and application in biocatalysis. FEMS Microbiol Rev 26: 73–81. doi: 10.1111/j.1574-6976.2002.tb00599.x
|
| [71] | Krastanova I, Guarnaccia C, Zahariev S, et al. (2005) Heterologous expression, purification, crystallization, X-ray analysis and phasing of the acetyl xylan esterase from Bacillus pumilus. BBA 1748: 222–230. |
| [72] |
Wong DWS (2006) Feruloyl esterase, a key enzyme in biomass degradation. Appl Biochem Biotechnol 133: 87–111. doi: 10.1385/ABAB:133:2:87
|
| [73] |
Dilokpimol A, Mäkelä MR, Aguilarpontes MV, et al. (2016) Diversity of fungal feruloyl esterases: Updated phylogenetic classification, properties, and industrial applications. Biotechnol Biofuels 9: 231. doi: 10.1186/s13068-016-0651-6
|
| [74] |
Faulds CB, Mandalari G, Curto RBL, et al. (2006) Synergy between xylanases from glycoside hydrolase family 10 and 11 and a feruloyl esterase in the release of phenolic acids from cereal arabinoxylan. Appl Microbiol Biotechnol 71: 622–629. doi: 10.1007/s00253-005-0184-6
|
| [75] | Wong DWS, Chan VJ, Liao H, et al. (2013) Cloning of a novel feruloyl esterase gene from rumen microbial metagenome and enzyme characterization in synergism with endoxylanases. J Ind Microbiol Biotechnol 40: 287–295. |
| [76] |
Pérezrodríguez N, Moreira CD, Torrado AA, et al. (2016) Feruloyl esterase production by Aspergillus terreus CECT 2808 and subsequent application to enzymatic hydrolysis. Enzyme Microb Tech 91: 52–58. doi: 10.1016/j.enzmictec.2016.05.011
|
| [77] |
Oliveira DM, Salvador VH, Mota TR, et al. (2016) Feruloyl esterase from Aspergillus clavatus improves xylan hydrolysis of sugarcane bagasse. AIMS Bioeng 4: 1–11. doi: 10.3934/bioeng.2017.1.1
|
| [78] | Garg G, Singh A, Kaur A, et al. (2016) Microbial pectinases: An ecofriendly tool of nature for industries. Biotech 6: 47. |
| [79] | Kant S, Vohra A, Gupta R (2013) Purification and physicochemical properties of polygalacturonase from Aspergillus niger MTCC 3323. Protein Expres Purif 87: 11–16. |
| [80] |
Chen Q, Jin Y, Zhang G, et al. (2012) Improving production of bioethanol from duckweed (Landoltia punctata) by pectinase pretreatment. Energies 5: 3019–3032. doi: 10.3390/en5083019
|
| [81] |
Kataria R, Ghosh S (2011) Saccharification of Kans grass using enzyme mixture from Trichoderma reesei for bioethanol production. Bioresource Technol 102: 9970–9975. doi: 10.1016/j.biortech.2011.08.023
|
| [82] | Oberoi HS, Vadlani PV, Nanjundaswamy A, et al. (2011) Enhanced ethanol production from Kinnow mandarin (Citrus reticulata) waste via a statistically optimized simultaneous saccharification and fermentation process. Bioresource Technol 102: 1593–1601. |
| [83] | Shanmugam VK, Gopalakrishnan D (2016) Screening and partial purification for the production of ligninase enzyme from the fungal isolate Trichosporon asahii. Braz Arch Biol Technol 59: e160220. |
| [84] |
Vrsanska M, Voberkova S, Langer V, et al. (2016) Induction of laccase, lignin peroxidase and manganese peroxidase activities in white-rot fungi using copper complexes. Molecules 21: E1553. doi: 10.3390/molecules21111553
|
| [85] | Pan L, Zhao H, Yu Q (2017) miR397/Laccase gene mediated network improves tolerance to fenoxaprop-P-ethyl in Beckmannia syzigachne and Oryza sativa. Front Plant Sci 23: 879. |
| [86] |
Kameshwar AKS, Qin W (2016) Recent developments in using advanced sequencing technologies for the genomic studies of lignin and cellulose degrading microorganisms. Int J Biol Sci 12: 156–171. doi: 10.7150/ijbs.13537
|
| [87] |
Rastogi M, Shrivastava S (2017) Recent advances in second generation bioethanol production: An insight to pretreatment, saccharification and fermentation processes. Renew Sust Energ Rev 80: 330–340. doi: 10.1016/j.rser.2017.05.225
|
| [88] |
Huang R, Su R, Qi W, et al. (2011) Bioconversion of lignocellulose into bioethanol process intensification and mechanism research. BioEnerg Res 4: 225–245. doi: 10.1007/s12155-011-9125-7
|
| [89] |
Lygin AV, Upton J, Dohleman FG, et al. (2011) Composition of cell wall phenolics and polysaccharides of the potential bioenergy crop-Miscanthus. Global Change Biol Bioenergy 3: 333–345. doi: 10.1111/j.1757-1707.2011.01091.x
|
| [90] |
Wei Y, Li X, Yu L, et al. (2015) Mesophilic anaerobic co-digestion of cattle manure and corn stover with biological and chemical pretreatment. Bioresource Technol 198: 431–436. doi: 10.1016/j.biortech.2015.09.035
|
| [91] |
Balat M, Balat H (2009) Recent trends in global production and utilization of bio-ethanol fuel. Appl Energ 86: 2273–2282. doi: 10.1016/j.apenergy.2009.03.015
|
| [92] | Cianchetta S, Maggio BD, Burzi PL, et al. (2014) Evaluation of selected white-rot fungal isolates for improving the sugar yield from wheat straw. Appl Biochem Biotechnol 173: 609–623. |
| [93] |
Behera S, Arora R, Nandhagopal N, et al. (2014) Importance of chemical pretreatment for bioconversion of lignocellulosic biomass. Renew Sust Energ Rev 36: 91–106. doi: 10.1016/j.rser.2014.04.047
|
| [94] |
Safari A, Karimi K, Shafiei M (2017) Dilute alkali pretreatment of softwood pine: A biorefinery approach. Bioresource Technol 234: 67–76. doi: 10.1016/j.biortech.2017.03.030
|
| [95] |
Rabemanolontsoa H, Saka S (2016) Various pretreatments of lignocellulosics. Bioresource Technol 199: 83–91. doi: 10.1016/j.biortech.2015.08.029
|
| [96] | Zhu Z, Simister R, Bird S, et al. (2015) Microwave assisted acid and alkali pretreatment of Miscanthus biomass for biorefineries. AIMS Bioeng 2: 446–468. |
| [97] |
Tang C, Shan J, Chen Y, et al. (2017) Organic amine catalytic organosolv pretreatment of corn stover for enzymatic saccharification and high-quality lignin. Bioresource Technol 232: 222–228. doi: 10.1016/j.biortech.2017.02.041
|
| [98] |
Silva ASD, Inoue H, Endo T, et al. (2010) Milling pretreatment of sugarcane bagasse and straw for enzymatic hydrolysis and ethanol fermentation. Bioresource Technol 101: 7402–7409. doi: 10.1016/j.biortech.2010.05.008
|
| [99] |
Chemat S, Lagha A, AitAmar H, et al. (2004) Comparison of conventional and ultrasound-assisted extraction of carvone and limonene from caraway seeds. Flavour Frag J 19: 188–195. doi: 10.1002/ffj.1339
|
| [100] | Sipponen MH, Rahikainen J, Leskinen T, et al. (2017) Structural changes of lignin in biorefinery pretreatments. Nord Pulp Pap Res J 32: 547–568. |
| [101] |
Fang H, Zhao C, Song XY, et al. (2013) Enhanced cellulolytic enzyme production by the synergism between Trichoderma reesei RUT-30 and Aspergillus niger NL02 and by the addition of surfactants. Biotechnol Bioprocess Eng 18: 390–398. doi: 10.1007/s12257-012-0562-8
|
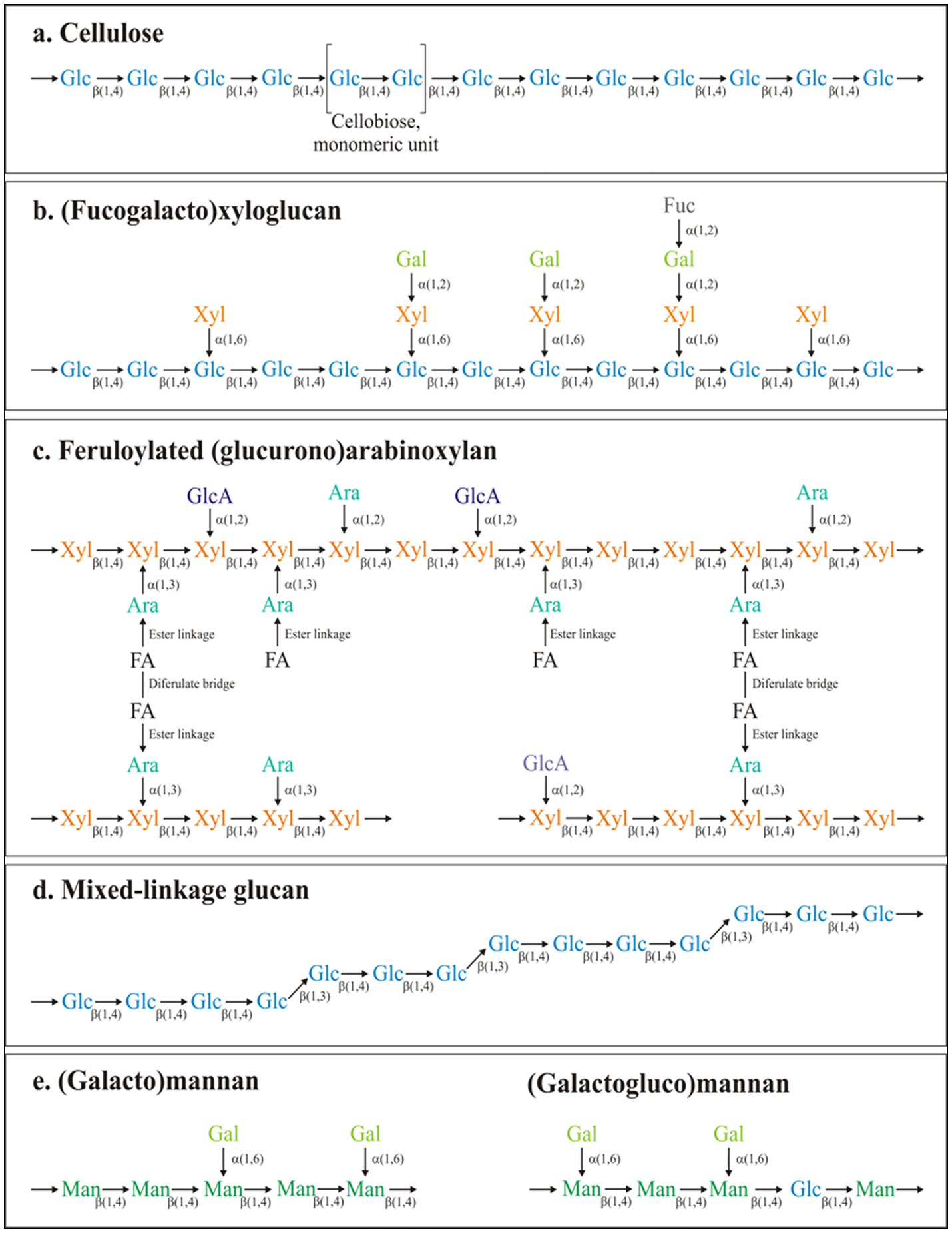
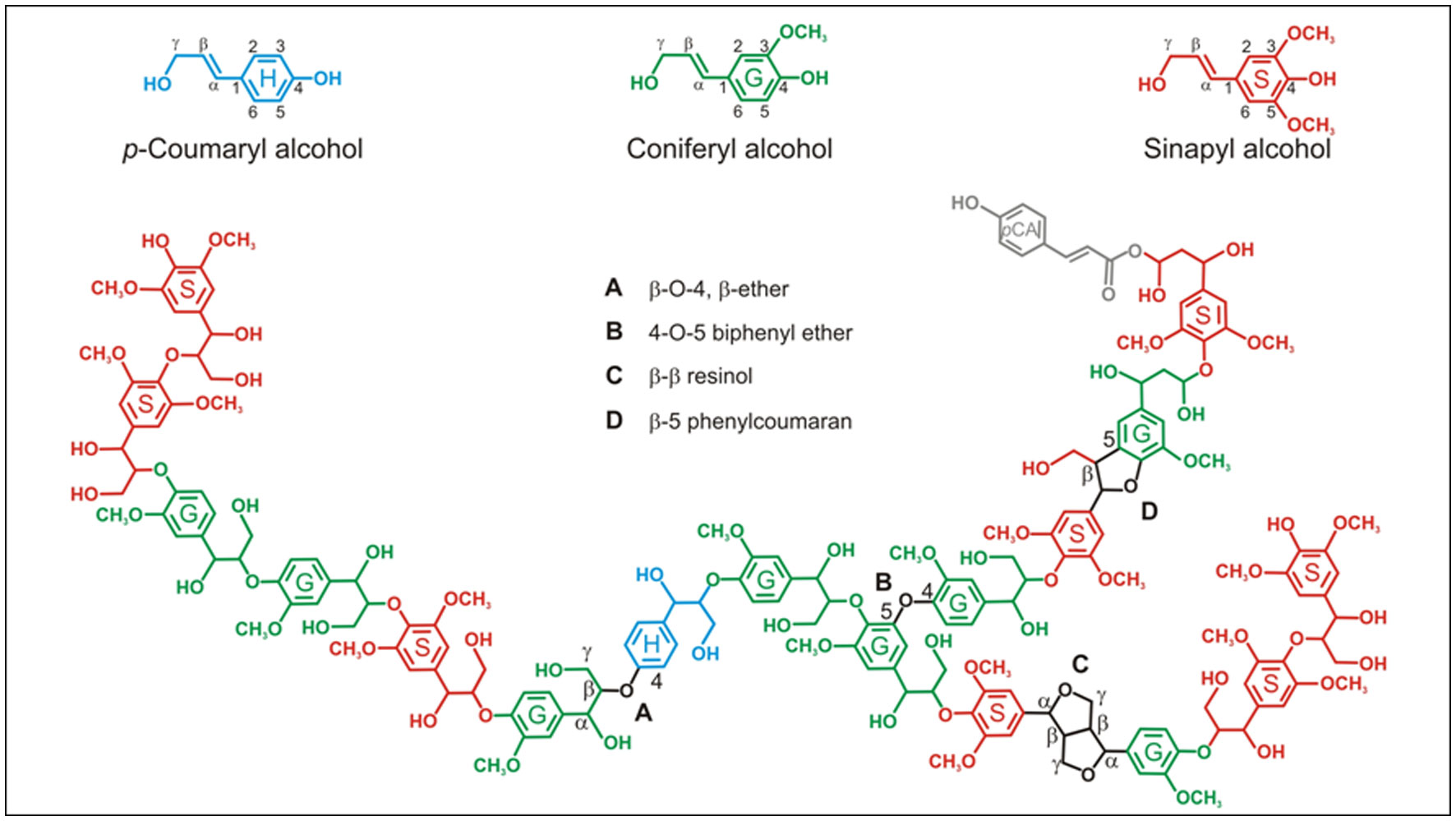
Figures(2) / Tables(1)

Thatiane Rodrigues Mota, Dyoni Matias de Oliveira, Rogério Marchiosi, Osvaldo Ferrarese-Filho, Wanderley Dantas dos Santos. Plant cell wall composition and enzymatic deconstruction[J]. AIMS Bioengineering, 2018, 5(1): 63-77. doi: 10.3934/bioeng.2018.1.63







 DownLoad:
DownLoad: