Citation: Dumisani Chirambo. Increasing the value of climate finance in an uncertain environment: Diaspora financial resources as a source of climate finance for Sub-Saharan Africa[J]. AIMS Environmental Science, 2017, 4(6): 730-742. doi: 10.3934/environsci.2017.6.730

| [1] |
Lau LC, Lee KT, Mohamed AR (2012) Global warming mitigation and renewable energy policy development from the Kyoto Protocol to the Copenhagen Accord-A comment. Renew Sust Energ Rev 16: 5280-5284. doi: 10.1016/j.rser.2012.04.006

|
| [2] |
Cobbinah PB, Erdiaw-Kwasie MO, Amoateng P (2015) Africa's urbanisation: implications for sustainable development. Cities 47: 62-72. doi: 10.1016/j.cities.2015.03.013
|
| [3] |
de Oliveira JAP (2009) The implementation of climate change related policies at the subnational level: An analysis of three countries. Habitat Int 33: 253-259. doi: 10.1016/j.habitatint.2008.10.006
|
| [4] |
Abadie LM, Galarraga I, Rübbelke D (2013) An analysis of the causes of the mitigation bias in international climate finance. Mitig Adapt Strat GL 18: 943-955. doi: 10.1007/s11027-012-9401-7
|
| [5] |
Morton TA, Rabinovich A, Marshall D, et al. (2011) The future that may (or may not) come: How framing changes responses to uncertainty in climate change communications. Global Environ Change 21: 103-109. doi: 10.1016/j.gloenvcha.2010.09.013
|
| [6] | Wilson RH, Smith TG, Urban resilience to climate change challenges in Africa. Working Paper No.4., The Robert S. Strauss Centre for International Security and Law, Texas. 2014. |
| [7] |
Scrieciu SS, Barker T, Ackerman F (2013) Pushing the boundaries of climate economics: critical issues to consider in climate policy analysis. Ecol Econ 85: 155-165. doi: 10.1016/j.ecolecon.2011.10.016
|
| [8] |
Hu Y, Monroy CR (2012) Chinese energy and climate policies after Durban: Save the Kyoto Protocol. Renew Sust Energ Rev 16: 3243-3250. doi: 10.1016/j.rser.2012.02.048
|
| [9] |
Rong F (2010) Understanding developing country stances on post-2012 climate change negotiations: Comparative analysis of Brazil, China, India, Mexico, and South Africa. Energy Policy 38: 4582-4591. doi: 10.1016/j.enpol.2010.04.014
|
| [10] | Howlett M (2014) Why are policy innovations rare and so often negative? Blame avoidance and problem denial in climate change policy-making. Global Environ Change 29: 395-403. |
| [11] |
Masini A, Menichetti E (2013) Investment decisions in the renewable energy sector: An analysis of nonfinancial drivers. Tech Forecast Soc Change 80: 510-524. doi: 10.1016/j.techfore.2012.08.003
|
| [12] | Hallegatte S, Bangalore M, Bonzanigo L, et al., Shock Waves: Managing the Impacts of Climate Change on Poverty. Climate Change and Development Series, World Bank, Washington, DC. 2016. |
| [13] |
Buurman J, Babovic V (2016) Adaptation Pathways and Real Options Analysis: An approach to deep uncertainty in climate change adaptation policies. Policy Soc 35: 137-150. doi: 10.1016/j.polsoc.2016.05.002
|
| [14] | Buchner B, Falconer A, Hervé-Mignucci M, et al., The Landscape of Climate Finance: A CPI report. Climate Policy Initiative, Venice, 2011. Available from: http://climatepolicyinitiative.org/wp-content/uploads/2011/10/The-Landscape-of-Climate-Finance-120120.pdf. |
| [15] | UN (United Nations), Transforming our world: the 2030 Agenda for Sustainable Development. United Nations, New York. 2015. |
| [16] |
Glemarec Y (2012) Financing off-grid sustainable energy access for the poor. Energy Policy 47: 87-93. doi: 10.1016/j.enpol.2012.03.032
|
| [17] | Yu Y (2014) Climate finance, Africa and China's role. Afr East-Asian Affair 1: 36-57. |
| [18] | Kato T, Ellis J, Pauw P, et al., Scaling up and replicating effective climate Finance interventions. Climate Change Expert Group Paper 2014 (1), OECD, Paris. 2014. |
| [19] | Mbeva K, Ochieng C, Atela J, et al., Intended Nationally Determined Contributions as a Means to Strengthening Africa's Engagement in International Climate Negotiations. Climate Resilient Economies Working Paper no. 001/2015, African Centre for Technology Studies (ACTS), ACTS Press, Nairobi. 2015. |
| [20] | Campillo G, Mullan M, Vallejo L, Climate Change Adaptation and Financial Protection: Synthesis of Key Findings from Colombia and Senegal. OECD Environment Working Papers No. 120, OECD Publishing, Paris. 2017. |
| [21] | World Bank, Africa's Pulse, Oct 2015, vol. 12, World Bank, Washington, DC. 2015. |
| [22] |
Eriksen S, Aldunce P, Bahinipati CS, et al. (2011) When not every response to climate change is a good one: Identifying principles for sustainable adaptation. Climate Dev 3:7-20. doi: 10.3763/cdev.2010.0060
|
| [23] |
Pasquini L, Cowling RM, Ziervogel G (2013) Facing the heat: Barriers to mainstreaming climate change adaptation in local government in the Western Cape Province, South Africa. Habitat Int 40: 225-232. doi: 10.1016/j.habitatint.2013.05.003
|
| [24] | AfDB (African Development Bank), Available from: http://www.afdb.org/en/cop/programme/climate-finance-for-africa/. |
| [25] |
Mitchell TD, Hulme M (1999) Predicting regional climate change: living with uncertainty. Prog Physl Geog 23: 57-78. doi: 10.1191/030913399672023346
|
| [26] | UNFCCC (United Nations Framework Convention on Climate Change), Report of the conference of the parties on its sixteenth session; addendum: part two: action taken by the conference of the parties at its sixteenth session. FCCC/CP/2010/7/Add.1. 2011. |
| [27] | Buchner B, Stadelmann M, Wilkinson J, et al., The Global Landscape of Climate Finance 2014. Climate Policy Initiative, Venice. 2014. |
| [28] |
Zhang W, Pan X (2016) Study on the demand of climate finance for developing countries based on submitted INDC. Adv Climate Change Res 7: 99-104. doi: 10.1016/j.accre.2016.05.002
|
| [29] |
Marwan NF, Kadir NA, Hussin A, et al. (2013) Export, aid, remittance and growth: Evidence from Sudan. Procedia Econ Financ 7: 3-10. doi: 10.1016/S2212-5671(13)00211-6
|
| [30] | Centre for Environmental Policy and Advocacy, Draft Position Paper: Towards Development of Climate Change Policy in Malawi. Centre for Environmental Policy and Advocacy, Blantyre. 2012. Available from: http://www.cepa.org.mw/documents/ECRP_documents/CEPA_Draft_Position_Paper_Climate_Change_Policy.pdf. |
| [31] | Bird N, Asante F, Bawakyillenuo S, et al., Public spending on climate change in Africa: Experiences from Ethiopia, Ghana, Tanzania and Uganda. Overseas Development Institute, London. 2016. |
| [32] | Niang I, Ruppel OC, Abdrabo MA, et al., Africa. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, pp. 1199-1265. 2014. |
| [33] |
Arezki R, Brückner M (2012) Rainfall financial development, and remittances: Evidence from Sub-Saharan Africa. J Int Econ 87: 377-385. doi: 10.1016/j.jinteco.2011.12.010
|
| [34] | Watkins K, Quattri M, Lost in intermediation: How excessive charges undermine the benefits of remittances for Africa, Overseas Development Institute, London. 2014. |
| [35] |
Kriegler E, O'Neill BC, Hallegatte S, et al. (2012) The need for and use of socio-economic scenarios for climate change analysis: A new approach based on shared socio-economic pathways. Global Environ Change 22: 807-822. doi: 10.1016/j.gloenvcha.2012.05.005
|
| [36] |
Anda J, Golub A, Strukova E (2009) Economics of climate change under uncertainty: Benefits of flexibility. Energy Policy 37: 1345-1355. doi: 10.1016/j.enpol.2008.11.034
|
| [37] |
Engels A, Hüther O, Schäfer M, et al. (2013) Public climate-change skepticism, energy preferences and political participation. Global Environ Change 23: 1018-1027. doi: 10.1016/j.gloenvcha.2013.05.008
|
| [38] |
Grafakos S, Flamos A, Oikonomou V, et al. (2010) Multi-criteria analysis weighting methodology to incorporate stakeholders' preferences in energy and climate policy interactions. Int J Energy Sec Manag 4: 434-461. doi: 10.1108/17506221011073851
|
| [39] |
Broughton EK, Brent AC, Haywood L (2012) Application of A Multi-Criteria Analysis Approach for Decision-Making in the Energy Sector: The Case of Concentrating Solar Power in South Africa. Energy Environ 23: 1221-1231. doi: 10.1260/0958-305X.23.8.1221
|
| [40] | Konidari P, Mavrakis D (2007) Multi-criteria evaluation of climate policy interactions. J Multi-criteria Decis Anal 14: 35-53. |
| [41] |
Kalim PFHB, Shah U (2011) A multi-criteria evaluation of policy instruments for climate change mitigation in the power generation sector of Trinidad and Tobago. Energy Policy 39: 6331-6343. doi: 10.1016/j.enpol.2011.07.034
|
| [42] | Röser F, Day T, Kurdziel M, After Paris: What is next for Intended Nationally Determined Contributions (INDCs)? International Partnership on Mitigation and MRV and New Climate Institute, 2016. |
| [43] | Day T, Röser F, Kurdziel M, Conditionality of Intended Nationally Determined Contributions (INDCs). International Partnership on Mitigation and MRV and New Climate Institute. 2016. |
| [44] | Hood C, Adkins L, Levina E, Overview of INDCs Submitted by 31 August 2015. Paper No. 2015(4). Climate Change Expert Group. Organisation for Economic Co-operation and Development (OECD), Paris. 2015. |
| [45] | Viljoen W (2013) Addressing climate change issue in eastern and southern Africa: EAC, COMESA, SADC and the TFTA. In Cape to Cairo: Exploring the Tripartite FTA agenda. Trade Law Centre and the Swedish Embassy, Nairobi. 1-35. |
| [46] |
Mohammed YS, Mustafa MW, Bashir N (2013) Status of renewable energy consumption and developmental challenges in Sub-Sahara Africa. Renew Sust Energ Rev 27: 453-463. doi: 10.1016/j.rser.2013.06.044
|
| [47] |
Javadi FS, Rismanchi B, Sarraf M, et al. (2013) Global policy of rural electrification. Renew Sust Energ Rev 19: 402-416. doi: 10.1016/j.rser.2012.11.053
|
| [48] | International Energy Agency (IEA), Boosting the Power Sector in Sub-Saharan Africa: China's involvement. IEA, Paris. 2016. |
| [49] | African Development Bank (AfDB), Sustainable Energy for All Africa Hub: Annual Report 2015–2016, African Development Bank, Abidjan, 2016. |
| [50] |
Winkelman AG, Moore MR (2011) Explaining the differential distribution of Clean Development Mechanism projects across host countries. Energy Policy 39: 1132-1143. doi: 10.1016/j.enpol.2010.11.036
|
| [51] | Burian M, Aren C, Sterk W, et al. Integrating Africa's Least Developed Countries into the Global Carbon Market: Analyzing CDM Implementation Barriers, Federal Ministry for the Environment, Nature Conservation and Nuclear Safety (BMU). 2011. |
| [52] | Arens C, Wang-Helmreich H, Hodes GS, et al., Assessing Support Activities by International Donors for CDM Development in Sub-Saharan Africa with Focus on Selected Least Developed Countries, Federal Ministry for the Environment, Nature Conservation and Nuclear Safety (BMU). 2011. |
| [53] |
Afful-Koomson T (2015) The Green Climate Fund in Africa: what should be different? Climate Dev 7: 367-379. doi: 10.1080/17565529.2014.951015
|
| [54] |
Greening LA, Bernow S (2004) Design of coordinated energy and environmental policies: use of multi-criteria decision-making. Energy Policy 32: 721-735. doi: 10.1016/j.enpol.2003.08.017
|
| [55] |
McKibbin WJ, Wilcoxen PJ (2009) Uncertainty and climate change policy design. J Policy Model 31: 463-477. doi: 10.1016/j.jpolmod.2008.12.001
|
| [56] |
Ha S, Hale T, Ogden P (2016) Climate finance in and between developing countries: an emerging opportunity to build on. Global Policy 7: 102-108. doi: 10.1111/1758-5899.12293
|
| [57] | Weigel M, China's Climate Change South-South Cooperation: Track Record and Future Direction, United Nations Development Programme in China, Beijing. 2016. |
| [58] | Chirambo D (2017) Enhancing Climate Change Resilience through Microfinance: Redefining the Climate Finance Paradigm to Promote Inclusive Growth in Africa. J Dev Soc 33: 1-24. |
| [59] | Wibeck V (2014) Enhancing learning, communication and public engagement about climate change-some lessons from recent literature. Environ Edu Res 20: 387-411. |
| [60] |
Nisbet MC (2009) Communicating Climate Change: Why Frames Matter for Public Engagement. Environ Sci Policy Sustain Dev 51: 12-23. doi: 10.3200/ENVT.51.2.12-23
|
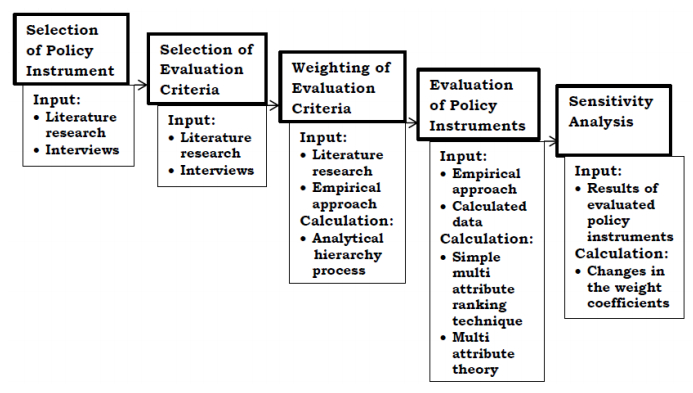
Figures(1) / Tables(1)

Dumisani Chirambo. Increasing the value of climate finance in an uncertain environment: Diaspora financial resources as a source of climate finance for Sub-Saharan Africa[J]. AIMS Environmental Science, 2017, 4(6): 730-742. doi: 10.3934/environsci.2017.6.730







 DownLoad:
DownLoad: