Citation: Nikhita Bolar, Aline Verstraeten, Lut Van Laer, Bart Loeys. Molecular insights into bicuspid aortic valve development and the associated aortopathy[J]. AIMS Molecular Science, 2017, 4(4): 478-508. doi: 10.3934/molsci.2017.4.478

| [1] |
Kelly RG (2012) The second heart field. Curr Top Dev Biol 100: 33-65. doi: 10.1016/B978-0-12-387786-4.00002-6

|
| [2] |
Buckingham M, Meilhac S, Zaffran S (2005) Building the mammalian heart from two sources of myocardial cells. Nat Rev Genet 6: 826-835. doi: 10.1038/nrg1710
|
| [3] | Kelly RG, Buckingham ME, Moorman AF (2014) Heart fields and cardiac morphogenesis. Cold Spring Harb Perspect Med 4. |
| [4] |
Yang YP, Li HR, Cao XM, et al. (2013) Second heart field and the development of the outflow tract in human embryonic heart. Dev Growth Differ 55: 359-367. doi: 10.1111/dgd.12050
|
| [5] |
Zaffran S, Frasch M (2002) Early signals in cardiac development. Circ Res 91: 457-469. doi: 10.1161/01.RES.0000034152.74523.A8
|
| [6] |
Srivastava D (2006) Making or breaking the heart: from lineage determination to morphogenesis. Cell 126: 1037-1048. doi: 10.1016/j.cell.2006.09.003
|
| [7] |
Graham A (2003) The neural crest. Curr Biol 13: R381-384. doi: 10.1016/S0960-9822(03)00315-4
|
| [8] |
van den Hoff MJ, Moorman AF (2000) Cardiac neural crest: the holy grail of cardiac abnormalities? Cardiovasc Res 47: 212-216. doi: 10.1016/S0008-6363(00)00127-9
|
| [9] |
Kirby ML, Hutson MR (2010) Factors controlling cardiac neural crest cell migration. Cell Adh Migr 4: 609-621. doi: 10.4161/cam.4.4.13489
|
| [10] |
Kirby ML, Waldo KL (1995) Neural crest and cardiovascular patterning. Circ Res 77: 211-215. doi: 10.1161/01.RES.77.2.211
|
| [11] |
Hutson MR, Kirby ML (2003) Neural crest and cardiovascular development: a 20-year perspective. Birth Defects Res C Embryo Today 69: 2-13. doi: 10.1002/bdrc.10002
|
| [12] |
Combs MD, Yutzey KE (2009) Heart valve development: regulatory networks in development and disease. Circ Res 105: 408-421. doi: 10.1161/CIRCRESAHA.109.201566
|
| [13] |
Lin CJ, Lin CY, Chen CH, et al. (2012) Partitioning the heart: mechanisms of cardiac septation and valve development. Development 139: 3277-3299. doi: 10.1242/dev.063495
|
| [14] | Feiner L, Webber AL, Brown CB, et al. (2001) Targeted disruption of semaphorin 3C leads to persistent truncus arteriosus and aortic arch interruption. Development 128: 3061-3070. |
| [15] | Brown CB, Feiner L, Lu MM, et al. (2001) PlexinA2 and semaphorin signaling during cardiac neural crest development. Development 128: 3071-3080. |
| [16] |
Toyofuku T, Yoshida J, Sugimoto T, et al. (2008) Repulsive and attractive semaphorins cooperate to direct the navigation of cardiac neural crest cells. Dev Biol 321: 251-262. doi: 10.1016/j.ydbio.2008.06.028
|
| [17] |
Verzi MP, McCulley DJ, De Val S, et al. (2005) The right ventricle, outflow tract, and ventricular septum comprise a restricted expression domain within the secondary/anterior heart field. Dev Biol 287: 134-145. doi: 10.1016/j.ydbio.2005.08.041
|
| [18] | Clouthier DE, Hosoda K, Richardson JA, et al. (1998) Cranial and cardiac neural crest defects in endothelin-A receptor-deficient mice. Development 125: 813-824. |
| [19] |
Koenig SN, Bosse KM, Nadorlik HA, et al. (2015) Evidence of Aortopathy in Mice with Haploinsufficiency of in -Null Background. J Cardiovasc Dev Dis 2: 17-30. doi: 10.3390/jcdd2010017
|
| [20] |
Feng Q, Song W, Lu X, et al. (2002) Development of heart failure and congenital septal defects in mice lacking endothelial nitric oxide synthase. Circulation 106: 873-879. doi: 10.1161/01.CIR.0000024114.82981.EA
|
| [21] |
Mommersteeg MT, Yeh ML, Parnavelas JG, et al. (2015) Disrupted Slit-Robo signalling results in membranous ventricular septum defects and bicuspid aortic valves. Cardiovasc Res 106: 55-66. doi: 10.1093/cvr/cvv040
|
| [22] |
Li P, Pashmforoush M, Sucov HM (2010) Retinoic acid regulates differentiation of the secondary heart field and TGFbeta-mediated outflow tract septation. Dev Cell 18: 480-485. doi: 10.1016/j.devcel.2009.12.019
|
| [23] |
Gruber PJ, Kubalak SW, Pexieder T, et al. (1996) RXR alpha deficiency confers genetic susceptibility for aortic sac, conotruncal, atrioventricular cushion, and ventricular muscle defects in mice. J Clin Invest 98: 1332-1343. doi: 10.1172/JCI118920
|
| [24] |
Zhou B, Wu B, Tompkins KL, et al. (2005) Characterization of Nfatc1 regulation identifies an enhancer required for gene expression that is specific to pro-valve endocardial cells in the developing heart. Development 132: 1137-1146. doi: 10.1242/dev.01640
|
| [25] | Bruneau BG (2013) Signaling and transcriptional networks in heart development and regeneration. Cold Spring Harb Perspect Biol 5: a008292. |
| [26] |
Lin CY, Lin CJ, Chen CH, et al. (2012) The secondary heart field is a new site of calcineurin/Nfatc1 signaling for semilunar valve development. J Mol Cell Cardiol 52: 1096-1102. doi: 10.1016/j.yjmcc.2012.01.013
|
| [27] | High FA, Jain R, Stoller JZ, et al. (2009) Murine Jagged1/Notch signaling in the second heart field orchestrates Fgf8 expression and tissue-tissue interactions during outflow tract development. J Clin Invest 119: 1986-1996. |
| [28] |
High FA, Zhang M, Proweller A, et al. (2007) An essential role for Notch in neural crest during cardiovascular development and smooth muscle differentiation. J Clin Invest 117: 353-363. doi: 10.1172/JCI30070
|
| [29] |
Chakraborty S, Combs MD, Yutzey KE (2010) Transcriptional regulation of heart valve progenitor cells. Pediatr Cardiol 31: 414-421. doi: 10.1007/s00246-009-9616-x
|
| [30] |
Shirai M, Imanaka-Yoshida K, Schneider MD, et al. (2009) T-box 2, a mediator of Bmp-Smad signaling, induced hyaluronan synthase2 and Tgfbeta2 expression and endocardial cushion formation. Proc Natl Acad Sci USA 106: 18604-18609. doi: 10.1073/pnas.0900635106
|
| [31] |
Chang CP, Neilson JR, Bayle JH, et al. (2004) A field of myocardial-endocardial NFAT signaling underlies heart valve morphogenesis. Cell 118: 649-663. doi: 10.1016/j.cell.2004.08.010
|
| [32] |
Schaefer BM, Lewin MB, Stout KK, et al. (2008) The bicuspid aortic valve: an integrated phenotypic classification of leaflet morphology and aortic root shape. Heart 94: 1634-1638. doi: 10.1136/hrt.2007.132092
|
| [33] |
Sievers HH, Schmidtke C (2007) A classification system for the bicuspid aortic valve from 304 surgical specimens. J Thorac Cardiovasc Surg 133: 1226-1233. doi: 10.1016/j.jtcvs.2007.01.039
|
| [34] |
Carro A, Teixido-Tura G, Evangelista A (2012) Aortic dilatation in bicuspid aortic valve disease. Rev Esp Cardiol 65: 977-981. doi: 10.1016/j.recesp.2012.04.025
|
| [35] | Girdauskas E, Borger MA, Secknus MA, et al. (2011) Is aortopathy in bicuspid aortic valve disease a congenital defect or a result of abnormal hemodynamics? A critical reappraisal of a one-sided argument. Eur J Cardiothorac Surg 39: 809-814. |
| [36] |
Jiao J, Xiong W, Wang L, et al. (2016) Differentiation defect in neural crest-derived smooth muscle cells in patients with aortopathy associated with bicuspid aortic valves. EBioMedicine 10: 282-290. doi: 10.1016/j.ebiom.2016.06.045
|
| [37] |
Loeys BL, Chen J, Neptune ER, et al. (2005) A syndrome of altered cardiovascular, craniofacial, neurocognitive and skeletal development caused by mutations in TGFBR1 or TGFBR2. Nat Genet 37: 275-281. doi: 10.1038/ng1511
|
| [38] | Kappetein AP, Gittenberger-de Groot AC, Zwinderman AH, et al. (1991) The neural crest as a possible pathogenetic factor in coarctation of the aorta and bicuspid aortic valve. J Thorac Cardiovasc Surg 102: 830-836. |
| [39] |
Martinez-Vargas J, Ventura J, Machuca A, et al. (2017) Cardiac, mandibular and thymic phenotypical association indicates that cranial neural crest underlies bicuspid aortic valve formation in hamsters. PLoS One 12: e0183556. doi: 10.1371/journal.pone.0183556
|
| [40] |
Van Hemelrijk C, Renard M, Loeys B (2010) The Loeys-Dietz syndrome: an update for the clinician. Curr Opin Cardiol 25: 546-551. doi: 10.1097/HCO.0b013e32833f0220
|
| [41] |
van de Laar IM, van der Linde D, Oei EH, et al. (2012) Phenotypic spectrum of the SMAD3- related aneurysms-osteoarthritis syndrome. J Med Genet 49: 47-57. doi: 10.1136/jmedgenet-2011-100382
|
| [42] |
Michelena HI, Khanna AD, Mahoney D, et al. (2011) Incidence of aortic complications in patients with bicuspid aortic valves. JAMA 306: 1104-1112. doi: 10.1001/jama.2011.1286
|
| [43] |
Cripe L, Andelfinger G, Martin LJ, et al. (2004) Bicuspid aortic valve is heritable. J Am Coll Cardiol 44: 138-143. doi: 10.1016/j.jacc.2004.03.050
|
| [44] |
Kang JW, Song HG, Yang DH, et al. (2013) Association between bicuspid aortic valve phenotype and patterns of valvular dysfunction and bicuspid aortopathy: comprehensive evaluation using MDCT and echocardiography. JACC Cardiovasc Imaging 6: 150-161. doi: 10.1016/j.jcmg.2012.11.007
|
| [45] |
Sans-Coma V, Fernandez B, Duran AC, et al. (1996) Fusion of valve cushions as a key factor in the formation of congenital bicuspid aortic valves in Syrian hamsters. Anat Rec 244: 490-498. doi: 10.1002/(SICI)1097-0185(199604)244:4<490::AID-AR7>3.0.CO;2-Z
|
| [46] |
Fernandez B, Duran AC, Fernandez-Gallego T, et al. (2009) Bicuspid aortic valves with different spatial orientations of the leaflets are distinct etiological entities. J Am Coll Cardiol 54: 2312-2318. doi: 10.1016/j.jacc.2009.07.044
|
| [47] |
Mahadevia R, Barker AJ, Schnell S, et al. (2014) Bicuspid aortic cusp fusion morphology alters aortic three-dimensional outflow patterns, wall shear stress, and expression of aortopathy. Circulation 129: 673-682. doi: 10.1161/CIRCULATIONAHA.113.003026
|
| [48] | Girdauskas E, Disha K, Borger MA, et al. (2012) Relation of bicuspid aortic valve morphology to the dilatation pattern of the proximal aorta: focus on the transvalvular flow. Cardiol Res Pract 2012: 478259. |
| [49] | Robicsek F, Thubrikar MJ, Cook JW, et al. (2004) The congenitally bicuspid aortic valve: how does it function? Why does it fail? Ann Thorac Surg 77: 177-185. |
| [50] |
Della Corte A, Bancone C, Conti CA, et al. (2012) Restricted cusp motion in right-left type of bicuspid aortic valves: a new risk marker for aortopathy. J Thorac Cardiovasc Surg 144: 360-369. doi: 10.1016/j.jtcvs.2011.10.014
|
| [51] |
Hope MD, Hope TA, Crook SE, et al. (2011) 4D flow CMR in assessment of valve-related ascending aortic disease. JACC Cardiovasc Imaging 4: 781-787. doi: 10.1016/j.jcmg.2011.05.004
|
| [52] |
Hope MD, Wrenn J, Sigovan M, et al. (2012) Imaging biomarkers of aortic disease: increased growth rates with eccentric systolic flow. J Am Coll Cardiol 60: 356-357. doi: 10.1016/j.jacc.2012.01.072
|
| [53] |
Ikonomidis JS, Ruddy JM, Benton SM, et al. (2012) Aortic dilatation with bicuspid aortic valves: cusp fusion correlates to matrix metalloproteinases and inhibitors. Ann Thorac Surg 93: 457-463. doi: 10.1016/j.athoracsur.2011.09.057
|
| [54] |
Russo CF, Cannata A, Lanfranconi M, et al. (2008) Is aortic wall degeneration related to bicuspid aortic valve anatomy in patients with valvular disease? J Thorac Cardiovasc Surg 136: 937-942. doi: 10.1016/j.jtcvs.2007.11.072
|
| [55] |
Fazel SS, Mallidi HR, Lee RS, et al. (2008) The aortopathy of bicuspid aortic valve disease has distinctive patterns and usually involves the transverse aortic arch. J Thorac Cardiovasc Surg 135: 901-907. doi: 10.1016/j.jtcvs.2008.01.022
|
| [56] | McNally A, Madan A, Sucosky P (2017) Morphotype-Dependent Flow Characteristics in Bicuspid Aortic Valve Ascending Aortas: A Benchtop Particle Image Velocimetry Study. Front Physiol 8: 44. |
| [57] |
Dolan JM, Meng H, Singh S, et al. (2011) High fluid shear stress and spatial shear stress gradients affect endothelial proliferation, survival, and alignment. Ann Biomed Eng 39: 1620- 1631. doi: 10.1007/s10439-011-0267-8
|
| [58] |
Rabkin SW (2014) Differential expression of MMP-2, MMP-9 and TIMP proteins in thoracic aortic aneurysm - comparison with and without bicuspid aortic valve: a meta-analysis. Vasa 43: 433-442. doi: 10.1024/0301-1526/a000390
|
| [59] |
Ikonomidis JS, Ivey CR, Wheeler JB, et al. (2013) Plasma biomarkers for distinguishing etiologic subtypes of thoracic aortic aneurysm disease. J Thorac Cardiovasc Surg 145: 1326- 1333. doi: 10.1016/j.jtcvs.2012.12.027
|
| [60] | Mohamed SA, Radtke A, Saraei R, et al. (2012) Locally different endothelial nitric oxide synthase protein levels in ascending aortic aneurysms of bicuspid and tricuspid aortic valve. Cardiol Res Pract 2012: 165957. |
| [61] |
Grewal N, DeRuiter MC, Jongbloed MR, et al. (2014) Normal and abnormal development of the aortic wall and valve: correlation with clinical entities. Neth Heart J 22: 363-369. doi: 10.1007/s12471-014-0576-2
|
| [62] |
Aicher D, Urbich C, Zeiher A, et al. (2007) Endothelial nitric oxide synthase in bicuspid aortic valve disease. Ann Thorac Surg 83: 1290-1294. doi: 10.1016/j.athoracsur.2006.11.086
|
| [63] |
Grewal N, Gittenberger-de Groot AC, DeRuiter MC, et al. (2014) Bicuspid aortic valve: phosphorylation of c-Kit and downstream targets are prognostic for future aortopathy. Eur J Cardiothorac Surg 46: 831-839. doi: 10.1093/ejcts/ezu319
|
| [64] |
Gurvitz M, Chang RK, Drant S, et al. (2004) Frequency of aortic root dilation in children with a bicuspid aortic valve. Am J Cardiol 94: 1337-1340. doi: 10.1016/j.amjcard.2004.07.130
|
| [65] |
Fernandes S, Khairy P, Graham DA, et al. (2012) Bicuspid aortic valve and associated aortic dilation in the young. Heart 98: 1014-1019. doi: 10.1136/heartjnl-2012-301773
|
| [66] | Yasuda H, Nakatani S, Stugaard M, et al. (2003) Failure to prevent progressive dilation of ascending aorta by aortic valve replacement in patients with bicuspid aortic valve: comparison with tricuspid aortic valve. Circulation 108 Suppl 1: II291-294. |
| [67] |
Silberbach M (2009) Bicuspid aortic valve and thoracic aortic aneurysm: toward a unified theory. J Am Coll Cardiol 53: 2296-2297. doi: 10.1016/j.jacc.2009.03.028
|
| [68] |
Bunton TE, Biery NJ, Myers L, et al. (2001) Phenotypic alteration of vascular smooth muscle cells precedes elastolysis in a mouse model of Marfan syndrome. Circ Res 88: 37-43. doi: 10.1161/01.RES.88.1.37
|
| [69] |
Tadros TM, Klein MD, Shapira OM (2009) Ascending aortic dilatation associated with bicuspid aortic valve: pathophysiology, molecular biology, and clinical implications. Circulation 119: 880-890. doi: 10.1161/CIRCULATIONAHA.108.795401
|
| [70] |
Bonderman D, Gharehbaghi-Schnell E, Wollenek G, et al. (1999) Mechanisms underlying aortic dilatation in congenital aortic valve malformation. Circulation 99: 2138-2143. doi: 10.1161/01.CIR.99.16.2138
|
| [71] |
Nistri S, Grande-Allen J, Noale M, et al. (2008) Aortic elasticity and size in bicuspid aortic valve syndrome. Eur Heart J 29: 472-479. doi: 10.1093/eurheartj/ehm528
|
| [72] |
Fedak PW, de Sa MP, Verma S, et al. (2003) Vascular matrix remodeling in patients with bicuspid aortic valve malformations: implications for aortic dilatation. J Thorac Cardiovasc Surg 126: 797-806. doi: 10.1016/S0022-5223(03)00398-2
|
| [73] | Nataatmadja M, West M, West J, et al. (2003) Abnormal extracellular matrix protein transport associated with increased apoptosis of vascular smooth muscle cells in marfan syndrome and bicuspid aortic valve thoracic aortic aneurysm. Circulation 108 Suppl 1: II329-334. |
| [74] |
Grewal N, Franken R, Mulder BJ, et al. (2016) Histopathology of aortic complications in bicuspid aortic valve versus Marfan syndrome: relevance for therapy? Heart Vessels 31: 795- 806. doi: 10.1007/s00380-015-0703-z
|
| [75] | Leme MP, David TE, Butany J, et al. (2003) Molecular evaluation of the great vessels of patients with bicuspid aortic valve disease. Braz J Cardiovasc Surg 18: 148-156. |
| [76] |
Rueda-Martinez C, Lamas O, Mataro MJ, et al. (2017) Fibrillin 2 is upregulated in the ascending aorta of patients with bicuspid aortic valve. Eur J Cardiothorac Surg 51: 104. doi: 10.1093/ejcts/ezw277
|
| [77] |
Siu SC, Silversides CK (2010) Bicuspid aortic valve disease. J Am Coll Cardiol 55: 2789-2800. doi: 10.1016/j.jacc.2009.12.068
|
| [78] |
Ward C (2000) Clinical significance of the bicuspid aortic valve. Heart 83: 81-85. doi: 10.1136/heart.83.1.81
|
| [79] |
Vallely MP, Semsarian C, Bannon PG (2008) Management of the ascending aorta in patients with bicuspid aortic valve disease. Heart Lung Circ 17: 357-363. doi: 10.1016/j.hlc.2008.01.007
|
| [80] |
Martin LJ, Ramachandran V, Cripe LH, et al. (2007) Evidence in favor of linkage to human chromosomal regions 18q, 5q and 13q for bicuspid aortic valve and associated cardiovascular malformations. Hum Genet 121: 275-284. doi: 10.1007/s00439-006-0316-9
|
| [81] |
Andelfinger G, Tapper AR, Welch RC, et al. (2002) KCNJ2 mutation results in Andersen syndrome with sex-specific cardiac and skeletal muscle phenotypes. Am J Hum Genet 71: 663-668. doi: 10.1086/342360
|
| [82] |
Garg V, Muth AN, Ransom JF, et al. (2005) Mutations in NOTCH1 cause aortic valve disease. Nature 437: 270-274. doi: 10.1038/nature03940
|
| [83] | Mohamed SA, Aherrahrou Z, Liptau H, et al. (2006) Novel missense mutations (p.T596M and p.P1797H) in NOTCH1 in patients with bicuspid aortic valve. Biochem Biophys Res Commun 345: 1460-1465. |
| [84] |
Acharya A, Hans CP, Koenig SN, et al. (2011) Inhibitory role of Notch1 in calcific aortic valve disease. PLoS One 6: e27743. doi: 10.1371/journal.pone.0027743
|
| [85] |
Quintero-Rivera F, Xi QJ, Keppler-Noreuil KM, et al. (2015) MATR3 disruption in human and mouse associated with bicuspid aortic valve, aortic coarctation and patent ductus arteriosus. Hum Mol Genet 24: 2375-2389. doi: 10.1093/hmg/ddv004
|
| [86] |
Moskowitz IP, Wang J, Peterson MA, et al. (2011) Transcription factor genes Smad4 and Gata4 cooperatively regulate cardiac valve development. Proc Natl Acad Sci USA 108: 4006-4011. doi: 10.1073/pnas.1019025108
|
| [87] |
Okubo A, Miyoshi O, Baba K, et al. (2004) A novel GATA4 mutation completely segregated with atrial septal defect in a large Japanese family. J Med Genet 41: e97. doi: 10.1136/jmg.2004.018895
|
| [88] |
Garg V, Kathiriya IS, Barnes R, et al. (2003) GATA4 mutations cause human congenital heart defects and reveal an interaction with TBX5. Nature 424: 443-447. doi: 10.1038/nature01827
|
| [89] |
Maitra M, Schluterman MK, Nichols HA, et al. (2009) Interaction of Gata4 and Gata6 with Tbx5 is critical for normal cardiac development. Dev Biol 326: 368-377. doi: 10.1016/j.ydbio.2008.11.004
|
| [90] |
Shi LM, Tao JW, Qiu XB, et al. (2014) GATA5 loss-of-function mutations associated with congenital bicuspid aortic valve. Int J Mol Med 33: 1219-1226. doi: 10.3892/ijmm.2014.1700
|
| [91] |
Bonachea EM, Chang SW, Zender G, et al. (2014) Rare GATA5 sequence variants identified in individuals with bicuspid aortic valve. Pediatr Res 76: 211-216. doi: 10.1038/pr.2014.67
|
| [92] |
Padang R, Bagnall RD, Richmond DR, et al. (2012) Rare non-synonymous variations in the transcriptional activation domains of GATA5 in bicuspid aortic valve disease. J Mol Cell Cardiol 53: 277-281 doi: 10.1016/j.yjmcc.2012.05.009
|
| [93] | Qu XK, Qiu XB, Yuan F, et al. (2014) A novel NKX2.5 loss-of-function mutation associated with congenital bicuspid aortic valve. Am J Cardiol 114: 1891-1895. |
| [94] |
Ponticos M, Partridge T, Black CM, et al. (2004) Regulation of collagen type I in vascular smooth muscle cells by competition between Nkx2.5 and deltaEF1/ZEB1. Mol Cell Biol 24: 6151-6161. doi: 10.1128/MCB.24.14.6151-6161.2004
|
| [95] |
Lin X, Huo Z, Liu X, et al. (2010) A novel GATA6 mutation in patients with tetralogy of Fallot or atrial septal defect. J Hum Genet 55: 662-667. doi: 10.1038/jhg.2010.84
|
| [96] |
Lepore JJ, Mericko PA, Cheng L, et al. (2006) GATA-6 regulates semaphorin 3C and is required in cardiac neural crest for cardiovascular morphogenesis. J Clin Invest 116: 929-939. doi: 10.1172/JCI27363
|
| [97] |
Kodo K, Nishizawa T, Furutani M, et al. (2009) GATA6 mutations cause human cardiac outflow tract defects by disrupting semaphorin-plexin signaling. Proc Natl Acad Sci USA 106: 13933-13938. doi: 10.1073/pnas.0904744106
|
| [98] | Pepe G, Nistri S, Giusti B, et al. (2014) Identification of fibrillin 1 gene mutations in patients with bicuspid aortic valve (BAV) without Marfan syndrome. BMC Med Genet 15: 23. |
| [99] |
Pereira L, Andrikopoulos K, Tian J, et al. (1997) Targetting of the gene encoding fibrillin-1 recapitulates the vascular aspect of Marfan syndrome. Nat Genet 17: 218-222. doi: 10.1038/ng1097-218
|
| [100] |
Loeys BL, Schwarze U, Holm T, et al. (2006) Aneurysm syndromes caused by mutations in the TGF-beta receptor. N Engl J Med 355: 788-798. doi: 10.1056/NEJMoa055695
|
| [101] |
van de Laar IM, Oldenburg RA, Pals G, et al. (2011) Mutations in SMAD3 cause a syndromic form of aortic aneurysms and dissections with early-onset osteoarthritis. Nat Genet 43: 121-126. doi: 10.1038/ng.744
|
| [102] |
Bertoli-Avella AM, Gillis E, Morisaki H, et al. (2015) Mutations in a TGF-beta ligand, TGFB3, cause syndromic aortic aneurysms and dissections. J Am Coll Cardiol 65: 1324-1336. doi: 10.1016/j.jacc.2015.01.040
|
| [103] |
Lindsay ME, Schepers D, Bolar NA, et al. (2012) Loss-of-function mutations in TGFB2 cause a syndromic presentation of thoracic aortic aneurysm. Nat Genet 44: 922-927. doi: 10.1038/ng.2349
|
| [104] |
Girdauskas E, Schulz S, Borger MA, et al. (2011) Transforming growth factor-beta receptor type II mutation in a patient with bicuspid aortic valve disease and intraoperative aortic dissection. Ann Thorac Surg 91: e70-71. doi: 10.1016/j.athoracsur.2010.12.060
|
| [105] | Folkersen L, Wagsater D, Paloschi V, et al. (2011) Unraveling divergent gene expression profiles in bicuspid and tricuspid aortic valve patients with thoracic aortic dilatation: the ASAP study. Mol Med 17: 1365-1373. |
| [106] |
Tan HL, Glen E, Topf A, et al. (2012) Nonsynonymous variants in the SMAD6 gene predispose to congenital cardiovascular malformation. Hum Mutat 33: 720-727. doi: 10.1002/humu.22030
|
| [107] |
Gillis E, Kumar AA, Luyckx I, et al. (2017) Candidate Gene Resequencing in a Large Bicuspid Aortic Valve-Associated Thoracic Aortic Aneurysm Cohort: SMAD6 as an Important Contributor. Frontiers in Physiology 8: 400. doi: 10.3389/fphys.2017.00400
|
| [108] |
Preuss C, Capredon M, Wunnemann F, et al. (2016) Family Based Whole Exome Sequencing Reveals the Multifaceted Role of Notch Signaling in Congenital Heart Disease. PLoS Genet 12: e1006335. doi: 10.1371/journal.pgen.1006335
|
| [109] |
Guo DC, Pannu H, Tran-Fadulu V, et al. (2007) Mutations in smooth muscle alpha-actin (ACTA2) lead to thoracic aortic aneurysms and dissections. Nat Genet 39: 1488-1493. doi: 10.1038/ng.2007.6
|
| [110] | Nigam V, Sievers HH, Jensen BC, et al. (2010) Altered microRNAs in bicuspid aortic valve: a comparison between stenotic and insufficient valves. J Heart Valve Dis 19: 459-465. |
| [111] |
Yanagawa B, Lovren F, Pan Y, et al. (2012) miRNA-141 is a novel regulator of BMP-2-mediated calcification in aortic stenosis. J Thorac Cardiovasc Surg 144: 256-262. doi: 10.1016/j.jtcvs.2011.10.097
|
| [112] |
McKellar SH, Tester DJ, Yagubyan M, et al. (2007) Novel NOTCH1 mutations in patients with bicuspid aortic valve disease and thoracic aortic aneurysms. J Thorac Cardiovasc Surg 134: 290-296. doi: 10.1016/j.jtcvs.2007.02.041
|
| [113] | Foffa I, Ait Ali L, Panesi P, et al. (2013) Sequencing of NOTCH1, GATA5, TGFBR1 and TGFBR2 genes in familial cases of bicuspid aortic valve. BMC Med Genet 14: 44. |
| [114] |
Kerstjens-Frederikse WS, van de Laar IM, Vos YJ, et al. (2016) Cardiovascular malformations caused by NOTCH1 mutations do not keep left: data on 428 probands with left-sided CHD and their families. Genet Med 18: 914-923. doi: 10.1038/gim.2015.193
|
| [115] |
Yang B, Zhou W, Jiao J, et al. (2017) Protein-altering and regulatory genetic variants near GATA4 implicated in bicuspid aortic valve. Nat Commun 8: 15481. doi: 10.1038/ncomms15481
|
| [116] |
Misra C, Sachan N, McNally CR, et al. (2012) Congenital heart disease-causing Gata4 mutation displays functional deficits in vivo. PLoS Genet 8: e1002690. doi: 10.1371/journal.pgen.1002690
|
| [117] | Schott JJ, Benson DW, Basson CT, et al. (1998) Congenital heart disease caused by mutations in the transcription factor NKX2-5. Science 281: 108-111. |
| [118] |
Reamon-Buettner SM, Hecker H, Spanel-Borowski K, et al. (2004) Novel NKX2-5 mutations in diseased heart tissues of patients with cardiac malformations. Am J Pathol 164: 2117-2125. doi: 10.1016/S0002-9440(10)63770-4
|
| [119] |
De Luca A, Sarkozy A, Consoli F, et al. (2010) Familial transposition of the great arteries caused by multiple mutations in laterality genes. Heart 96: 673-677. doi: 10.1136/hrt.2009.181685
|
| [120] | Draus JM, Hauck MA, Goetsch M, et al. (2009) Investigation of somatic NKX2-5 mutations in congenital heart disease. J Med Genet 46: 115-122. |
| [121] |
Wooten EC, Iyer LK, Montefusco MC, et al. (2010) Application of gene network analysis techniques identifies AXIN1/PDIA2 and endoglin haplotypes associated with bicuspid aortic valve. PLoS One 5: e8830. doi: 10.1371/journal.pone.0008830
|
| [122] |
Martin M, Lorca R, Rozado J, et al. (2017) Bicuspid aortic valve syndrome: a multidisciplinary approach for a complex entity. J Thorac Dis 9: S454-S464. doi: 10.21037/jtd.2017.05.11
|
| [123] |
Guo DC, Gong L, Regalado ES, et al. (2015) MAT2A mutations predispose individuals to thoracic aortic aneurysms. Am J Hum Genet 96: 170-177. doi: 10.1016/j.ajhg.2014.11.015
|
| [124] |
Mohamed SA, Hanke T, Schlueter C, et al. (2005) Ubiquitin fusion degradation 1-like gene dysregulation in bicuspid aortic valve. J Thorac Cardiovasc Surg 130: 1531-1536. doi: 10.1016/j.jtcvs.2005.08.017
|
| [125] |
Dargis N, Lamontagne M, Gaudreault N, et al. (2016) Identification of Gender-Specific Genetic Variants in Patients With Bicuspid Aortic Valve. Am J Cardiol 117: 420-426. doi: 10.1016/j.amjcard.2015.10.058
|
| [126] | Malfait F, Symoens S, Coucke P, et al. (2006) Total absence of the alpha2(I) chain of collagen type I causes a rare form of Ehlers-Danlos syndrome with hypermobility and propensity to cardiac valvular problems. J Med Genet 43: e36. |
| [127] |
Tischfield MA, Bosley TM, Salih MA, et al. (2005) Homozygous HOXA1 mutations disrupt human brainstem, inner ear, cardiovascular and cognitive development. Nat Genet 37: 1035-1037. doi: 10.1038/ng1636
|
| [128] |
Jain R, Engleka KA, Rentschler SL, et al. (2011) Cardiac neural crest orchestrates remodeling and functional maturation of mouse semilunar valves. J Clin Invest 121: 422-430. doi: 10.1172/JCI44244
|
| [129] |
Garg V (2006) Molecular genetics of aortic valve disease. Curr Opin Cardiol 21: 180-184. doi: 10.1097/01.hco.0000221578.18254.70
|
| [130] |
Macatee TL, Hammond BP, Arenkiel BR, et al. (2003) Ablation of specific expression domains reveals discrete functions of ectoderm- and endoderm-derived FGF8 during cardiovascular and pharyngeal development. Development 130: 6361-6374. doi: 10.1242/dev.00850
|
| [131] |
Laforest B, Andelfinger G, Nemer M (2011) Loss of Gata5 in mice leads to bicuspid aortic valve. J Clin Invest 121: 2876-2887. doi: 10.1172/JCI44555
|
| [132] |
Lee TC, Zhao YD, Courtman DW, et al. (2000) Abnormal aortic valve development in mice lacking endothelial nitric oxide synthase. Circulation 101: 2345-2348. doi: 10.1161/01.CIR.101.20.2345
|
| [133] |
Laforest B, Nemer M (2011) GATA5 interacts with GATA4 and GATA6 in outflow tract development. Dev Biol 358: 368-378. doi: 10.1016/j.ydbio.2011.07.037
|
| [134] |
Timmerman LA, Grego-Bessa J, Raya A, et al. (2004) Notch promotes epithelial-mesenchymal transition during cardiac development and oncogenic transformation. Genes Dev 18: 99-115. doi: 10.1101/gad.276304
|
| [135] |
Biben C, Weber R, Kesteven S, et al. (2000) Cardiac septal and valvular dysmorphogenesis in mice heterozygous for mutations in the homeobox gene Nkx2-5. Circ Res 87: 888-895. doi: 10.1161/01.RES.87.10.888
|
| [136] |
Kaartinen V, Dudas M, Nagy A, et al. (2004) Cardiac outflow tract defects in mice lacking ALK2 in neural crest cells. Development 131: 3481-3490. doi: 10.1242/dev.01214
|
| [137] |
Thomas PS, Sridurongrit S, Ruiz-Lozano P, et al. (2012) Deficient signaling via Alk2 (Acvr1) leads to bicuspid aortic valve development. PLoS One 7: e35539. doi: 10.1371/journal.pone.0035539
|
| [138] |
Stottmann RW, Choi M, Mishina Y, et al. (2004) BMP receptor IA is required in mammalian neural crest cells for development of the cardiac outflow tract and ventricular myocardium. Development 131: 2205-2218. doi: 10.1242/dev.01086
|
| [139] |
Jia Q, McDill BW, Li SZ, et al. (2007) Smad signaling in the neural crest regulates cardiac outflow tract remodeling through cell autonomous and non-cell autonomous effects. Dev Biol 311: 172-184. doi: 10.1016/j.ydbio.2007.08.044
|
| [140] |
Cai X, Zhang W, Hu J, et al. (2013) Tbx20 acts upstream of Wnt signaling to regulate endocardial cushion formation and valve remodeling during mouse cardiogenesis. Development 140: 3176-3187. doi: 10.1242/dev.092502
|
| [141] |
Takeuchi JK, Mileikovskaia M, Koshiba-Takeuchi K, et al. (2005) Tbx20 dose-dependently regulates transcription factor networks required for mouse heart and motoneuron development. Development 132: 2463-2474. doi: 10.1242/dev.01827
|
| [142] |
Clouthier DE, Williams SC, Hammer RE, et al. (2003) Cell-autonomous and nonautonomous actions of endothelin-A receptor signaling in craniofacial and cardiovascular development. Dev Biol 261: 506-519. doi: 10.1016/S0012-1606(03)00128-3
|
| [143] |
Kurihara Y, Kurihara H, Oda H, et al. (1995) Aortic arch malformations and ventricular septal defect in mice deficient in endothelin-1. J Clin Invest 96: 293-300. doi: 10.1172/JCI118033
|
| [144] |
Yanagisawa H, Hammer RE, Richardson JA, et al. (2000) Disruption of ECE-1 and ECE-2 reveals a role for endothelin-converting enzyme-2 in murine cardiac development. J Clin Invest 105: 1373-1382. doi: 10.1172/JCI7447
|
| [145] | Yanagisawa H, Yanagisawa M, Kapur RP, et al. (1998) Dual genetic pathways of endothelin- mediated intercellular signaling revealed by targeted disruption of endothelin converting enzyme-1 gene. Development 125: 825-836. |
| [146] | Gitler AD, Zhu Y, Ismat FA, et al. (2003) Nf1 has an essential role in endothelial cells. Nat Genet 33: 75-79. |
| [147] |
Ranger AM, Grusby MJ, Hodge MR, et al. (1998) The transcription factor NF-ATc is essential for cardiac valve formation. Nature 392: 186-190. doi: 10.1038/32426
|
| [148] |
de la Pompa JL, Timmerman LA, Takimoto H, et al. (1998) Role of the NF-ATc transcription factor in morphogenesis of cardiac valves and septum. Nature 392: 182-186. doi: 10.1038/32419
|
| [149] |
Todorovic V, Frendewey D, Gutstein DE, et al. (2007) Long form of latent TGF-beta binding protein 1 (Ltbp1L) is essential for cardiac outflow tract septation and remodeling. Development 134: 3723-3732. doi: 10.1242/dev.008599
|
| [150] |
Todorovic V, Finnegan E, Freyer L, et al. (2011) Long form of latent TGF-beta binding protein 1 (Ltbp1L) regulates cardiac valve development. Dev Dyn 240: 176-187. doi: 10.1002/dvdy.22521
|
| [151] |
Escot S, Blavet C, Hartle S, et al. (2013) Misregulation of SDF1-CXCR4 signaling impairs early cardiac neural crest cell migration leading to conotruncal defects. Circ Res 113: 505-516. doi: 10.1161/CIRCRESAHA.113.301333
|
| [152] |
Yu S, Crawford D, Tsuchihashi T, et al. (2011) The chemokine receptor CXCR7 functions to 930 regulate cardiac valve remodeling. Dev Dyn 240: 384-393. doi: 10.1002/dvdy.22549
|
| [153] |
Thomas PS, Kim J, Nunez S, et al. (2010) Neural crest cell-specific deletion of Rac1 results in defective cell-matrix interactions and severe craniofacial and cardiovascular malformations. Dev Biol 340: 613-625. doi: 10.1016/j.ydbio.2010.02.021
|
| [154] | Shirai M, Osugi T, Koga H, et al. (2002) The Polycomb-group gene Rae28 sustains Nkx2.5/Csx expression and is essential for cardiac morphogenesis. J Clin Invest 110: 177-184. |
| [155] | Takihara Y, Tomotsune D, Shirai M, et al. (1997) Targeted disruption of the mouse homologue of the Drosophila polyhomeotic gene leads to altered anteroposterior patterning and neural crest defects. Development 124: 3673-3682. |
| [156] |
Gao Z, Kim GH, Mackinnon AC, et al. (2010) Ets1 is required for proper migration and differentiation of the cardiac neural crest. Development 137: 1543-1551. doi: 10.1242/dev.047696
|
| [157] |
Makki N, Capecchi MR (2012) Cardiovascular defects in a mouse model of HOXA1 syndrome. Hum Mol Genet 21: 26-31. doi: 10.1093/hmg/ddr434
|
| [158] |
Zhou J, Zhang M, Fang H, et al. (2009) The SWI/SNF chromatin remodeling complex regulates myocardin-induced smooth muscle-specific gene expression. Arterioscler Thromb Vasc Biol 29: 921-928. doi: 10.1161/ATVBAHA.109.187229
|
| [159] |
Zhang M, Fang H, Zhou J, et al. (2007) A novel role of Brg1 in the regulation of SRF/MRTFA- dependent smooth muscle-specific gene expression. J Biol Chem 282: 25708-25716. doi: 10.1074/jbc.M701925200
|
| [160] |
Zhang M, Chen M, Kim JR, et al. (2011) SWI/SNF complexes containing Brahma or Brahma-related gene 1 play distinct roles in smooth muscle development. Mol Cell Biol 31: 2618-2631. doi: 10.1128/MCB.01338-10
|
| [161] |
Theodoris CV, Li M, White MP, et al. (2015) Human disease modeling reveals integrated transcriptional and epigenetic mechanisms of NOTCH1 haploinsufficiency. Cell 160: 1072-1086. doi: 10.1016/j.cell.2015.02.035
|
| [162] |
Guarani V, Deflorian G, Franco CA, et al. (2011) Acetylation-dependent regulation of endothelial Notch signalling by the SIRT1 deacetylase. Nature 473: 234-238. doi: 10.1038/nature09917
|
| [163] | Yuan SM (2013) Congenital heart defects in Kabuki syndrome. Cardiol J 20: 121-124. |
| [164] |
Kim SJ, Cho SY, Maeng SH, et al. (2013) A novel MLL2 gene mutation in a Korean patient with Kabuki syndrome. Korean J Pediatr 56: 355-358. doi: 10.3345/kjp.2013.56.8.355
|
| [165] |
Kakita T, Hasegawa K, Morimoto T, et al. (1999) p300 protein as a coactivator of GATA-5 in the transcription of cardiac-restricted atrial natriuretic factor gene. J Biol Chem 274: 34096-34102. doi: 10.1074/jbc.274.48.34096
|
| [166] |
Stevens CA, Bhakta MG (1995) Cardiac abnormalities in the Rubinstein-Taybi syndrome. Am J Med Genet 59: 346-348. doi: 10.1002/ajmg.1320590313
|
| [167] | Kurtovic S, Paloschi V, Folkersen L, et al. (2011) Diverging alternative splicing fingerprints in the transforming growth factor-beta signaling pathway identified in thoracic aortic aneurysms. Mol Med 17: 665-675. |
| [168] |
Lamouille S, Xu J, Derynck R (2014) Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol 15: 178-196. doi: 10.1038/nrm3758
|
| [169] |
Brown CB, Boyer AS, Runyan RB, et al. (1996) Antibodies to the Type II TGFbeta receptor block cell activation and migration during atrioventricular cushion transformation in the heart. Dev Biol 174: 248-257. doi: 10.1006/dbio.1996.0070
|
| [170] |
Ramsdell AF, Markwald RR (1997) Induction of endocardial cushion tissue in the avian heart is regulated, in part, by TGFbeta-3-mediated autocrine signaling. Dev Biol 188: 64-74. doi: 10.1006/dbio.1997.8637
|
| [171] |
Boyer AS, Ayerinskas II, Vincent EB, et al. (1999) TGFbeta2 and TGFbeta3 have separate and sequential activities during epithelial-mesenchymal cell transformation in the embryonic heart. Dev Biol 208: 530-545. doi: 10.1006/dbio.1999.9211
|
| [172] |
Brown CB, Boyer AS, Runyan RB, et al. (1999) Requirement of type III TGF-beta receptor for endocardial cell transformation in the heart. Science 283: 2080-2082. doi: 10.1126/science.283.5410.2080
|
| [173] | Millan FA, Denhez F, Kondaiah P, et al. (1991) Embryonic gene expression patterns of TGF beta 1, beta 2 and beta 3 suggest different developmental functions in vivo. Development 111: 131-143. |
| [174] |
Nieto MA (2002) The snail superfamily of zinc-finger transcription factors. Nat Rev Mol Cell Biol 3: 155-166. doi: 10.1038/nrm757
|
| [175] |
Nieto MA, Sargent MG, Wilkinson DG, et al. (1994) Control of cell behavior during vertebrate development by Slug, a zinc finger gene. Science 264: 835-839. doi: 10.1126/science.7513443
|
| [176] |
Romano LA, Runyan RB (2000) Slug is an essential target of TGFbeta2 signaling in the developing chicken heart. Dev Biol 223: 91-102. doi: 10.1006/dbio.2000.9750
|
| [177] |
Carver EA, Jiang R, Lan Y, et al. (2001) The mouse snail gene encodes a key regulator of the epithelial-mesenchymal transition. Mol Cell Biol 21: 8184-8188. doi: 10.1128/MCB.21.23.8184-8188.2001
|
| [178] |
Liebner S, Cattelino A, Gallini R, et al. (2004) Beta-catenin is required for endothelial-mesenchymal transformation during heart cushion development in the mouse. J Cell Biol 166: 359-367. doi: 10.1083/jcb.200403050
|
| [179] |
Luna-Zurita L, Prados B, Grego-Bessa J, et al. (2010) Integration of a Notch-dependent mesenchymal gene program and Bmp2-driven cell invasiveness regulates murine cardiac valve formation. J Clin Invest 120: 3493-3507. doi: 10.1172/JCI42666
|
| [180] |
Galvin KM, Donovan MJ, Lynch CA, et al. (2000) A role for smad6 in development and homeostasis of the cardiovascular system. Nat Genet 24: 171-174. doi: 10.1038/72835
|
| [181] |
Takase M, Imamura T, Sampath TK, et al. (1998) Induction of Smad6 mRNA by bone morphogenetic proteins. Biochem Biophys Res Commun 244: 26-29. doi: 10.1006/bbrc.1998.8200
|
| [182] |
Corson LB, Yamanaka Y, Lai KM, et al. (2003) Spatial and temporal patterns of ERK signaling during mouse embryogenesis. Development 130: 4527-4537. doi: 10.1242/dev.00669
|
| [183] |
Newbern J, Zhong J, Wickramasinghe RS, et al. (2008) Mouse and human phenotypes indicate a critical conserved role for ERK2 signaling in neural crest development. Proc Natl Acad Sci USA 105: 17115-17120. doi: 10.1073/pnas.0805239105
|
| [184] |
Rivera-Feliciano J, Tabin CJ (2006) Bmp2 instructs cardiac progenitors to form the heart-1006 valve-inducing field. Dev Biol 295: 580-588. doi: 10.1016/j.ydbio.2006.03.043
|
| [185] |
Ma L, Lu MF, Schwartz RJ, et al. (2005) Bmp2 is essential for cardiac cushion epithelial-mesenchymal transition and myocardial patterning. Development 132: 5601-5611. doi: 10.1242/dev.02156
|
| [186] |
Sugi Y, Yamamura H, Okagawa H, et al. (2004) Bone morphogenetic protein-2 can mediate myocardial regulation of atrioventricular cushion mesenchymal cell formation in mice. Dev Biol 269: 505-518. doi: 10.1016/j.ydbio.2004.01.045
|
| [187] |
McCulley DJ, Kang JO, Martin JF, et al. (2008) BMP4 is required in the anterior heart field and its derivatives for endocardial cushion remodeling, outflow tract septation, and semilunar valve development. Dev Dyn 237: 3200-3209. doi: 10.1002/dvdy.21743
|
| [188] |
Kim RY, Robertson EJ, Solloway MJ (2001) Bmp6 and Bmp7 are required for cushion formation and septation in the developing mouse heart. Dev Biol 235: 449-466. doi: 10.1006/dbio.2001.0284
|
| [189] |
Delot EC, Bahamonde ME, Zhao M, et al. (2003) BMP signaling is required for septation of the outflow tract of the mammalian heart. Development 130: 209-220. doi: 10.1242/dev.00181
|
| [190] |
Beppu H, Malhotra R, Beppu Y, et al. (2009) BMP type II receptor regulates positioning of outflow tract and remodeling of atrioventricular cushion during cardiogenesis. Dev Biol 331: 167-175. doi: 10.1016/j.ydbio.2009.04.032
|
| [191] |
Gaussin V, Van de Putte T, Mishina Y, et al. (2002) Endocardial cushion and myocardial defects after cardiac myocyte-specific conditional deletion of the bone morphogenetic protein receptor ALK3. Proc Natl Acad Sci U S A 99: 2878-2883. doi: 10.1073/pnas.042390499
|
| [192] |
Radtke F, MacDonald HR, Tacchini-Cottier F (2013) Regulation of innate and adaptive immunity by Notch. Nat Rev Immunol 13: 427-437. doi: 10.1038/nri3445
|
| [193] |
Niessen K, Karsan A (2008) Notch signaling in cardiac development. Circ Res 102: 1169-1181. doi: 10.1161/CIRCRESAHA.108.174318
|
| [194] |
McDaniell R, Warthen DM, Sanchez-Lara PA, et al. (2006) NOTCH2 mutations cause Alagille syndrome, a heterogeneous disorder of the notch signaling pathway. Am J Hum Genet 79: 169-173. doi: 10.1086/505332
|
| [195] |
Warthen DM, Moore EC, Kamath BM, et al. (2006) Jagged1 (JAG1) mutations in Alagille syndrome: increasing the mutation detection rate. Hum Mutat 27: 436-443. doi: 10.1002/humu.20310
|
| [196] |
Donovan J, Kordylewska A, Jan YN, et al. (2002) Tetralogy of fallot and other congenital heart defects in Hey2 mutant mice. Curr Biol 12: 1605-1610. doi: 10.1016/S0960-9822(02)01149-1
|
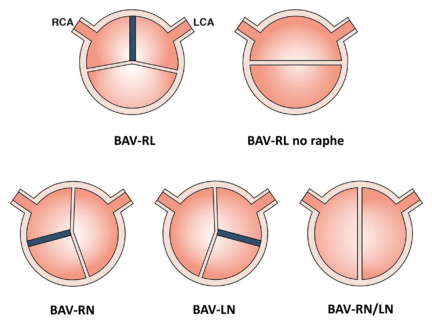
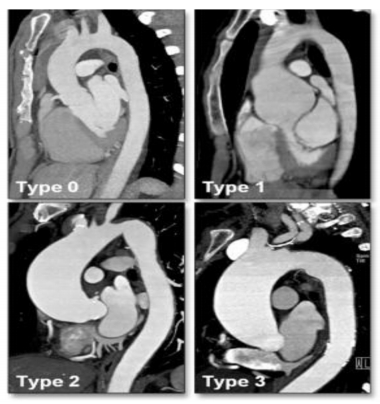
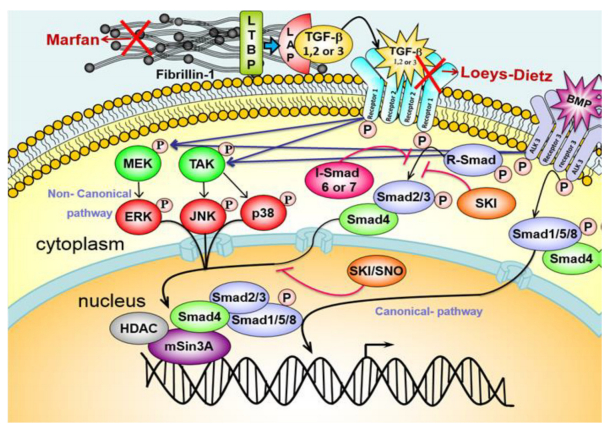
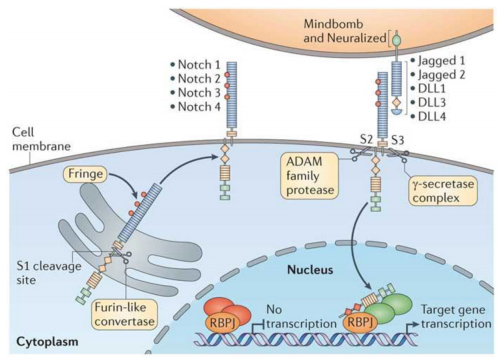
Figures(4) / Tables(2)

Nikhita Bolar, Aline Verstraeten, Lut Van Laer, Bart Loeys. Molecular insights into bicuspid aortic valve development and the associated aortopathy[J]. AIMS Molecular Science, 2017, 4(4): 478-508. doi: 10.3934/molsci.2017.4.478







 DownLoad:
DownLoad: