Citation: Mario P. Carante, Francesca Ballarini. Modelling cell death for cancer hadrontherapy[J]. AIMS Biophysics, 2017, 4(3): 465-490. doi: 10.3934/biophy.2017.3.465

| [1] |
Durante M, Loeffler JS (2010) Charged particles in radiation oncology. Nat Rev Clin Oncol 7: 37–43. doi: 10.1038/nrclinonc.2009.183

|
| [2] | Particle Therapy Co-Operative Group (PTCOG). Available from: http://www.ptcog.ch. |
| [3] |
Tommasino F, Scifoni E, Durante M (2015) New ions for therapy. Int J Particle Ther 2: 428–438. doi: 10.14338/IJPT-15-00027.1
|
| [4] | Weyrather WK (2004) Medical Applications of Accelerated Ions, In: Al-Khalili J, Roeckl E, The Euroschool Lectures on Physics with Exotic Beams, Berlin, Heidelberg: Springer Berlin Heidelberg, 469–490. |
| [5] | Molière G (1948) Theorie der streuung schneller geladener teilchen ii mehrfach-und vielfachstreuung. Z Naturforschung A 3: 78–97. |
| [6] |
Amaldi U, Kraft G (2005) Radiotherapy with beams of carbon ions. Rep Prog Phys 68: 1861–1882. doi: 10.1088/0034-4885/68/8/R04
|
| [7] | Linz U (2012) Physical and biological rationale for using ions in therapy, In: Ion Beam Therapy, Berlin, Heidelberg: Springer Berlin Heidelberg, 45–59. |
| [8] | Battistoni G, Bauer J, Boehlen TT, et al. (2016) The FLUKA code: An accurate simulation tool for particle therapy. Front Oncol 6: 116. |
| [9] |
Sato T, Niita K, Matsuda N, et al. (2014) Overview of the PHITS code and its application to medical physics. Prog Nucl Sci Technol 4: 879–882. doi: 10.15669/pnst.4.879
|
| [10] |
Allison J, Amaco K, Apostolakis J, et al. (2016) Recent developments in Geant4. Nucl Instrum Meth A 835: 186–225. doi: 10.1016/j.nima.2016.06.125
|
| [11] |
Scholz M, Kraft G (1994) Calculation of heavy ion inactivation probabilities based on track structure, X ray sensitivity and target size. Radiat Prot Dosim 52: 29–33. doi: 10.1093/oxfordjournals.rpd.a082156
|
| [12] |
Kase Y, Kanai T, Matsumoto Y, et al. (2006) Microdosimetric measurements and estimation of human cell survival for heavy-ion beams. Radiat Res 166: 629–638. doi: 10.1667/RR0536.1
|
| [13] |
Wouters BG, Lam GK, Oelfke U, et al. (1996) Measurements of relative biological effectiveness of the 70 MeV proton beam at TRIUMF using V79 cells and the high precision cell sorter assay. Radiat Res 146: 159–170. doi: 10.2307/3579588
|
| [14] |
Gerweck LE, Kozin SV (1999) Relative biological effectiveness of proton beams in clinical therapy. Radiother Oncol 50: 135–142. doi: 10.1016/S0167-8140(98)00092-9
|
| [15] |
Paganetti H (2002) Relative Biological Effectiveness (RBE) values for proton beam therapy. J Radiat Oncol Biol Phys 53: 407–421. doi: 10.1016/S0360-3016(02)02754-2
|
| [16] |
Paganetti H, van Luijk P (2013) Biological considerations when comparing proton therapy and photon therapy. Semin Radiat Oncol 23: 77–87. doi: 10.1016/j.semradonc.2012.11.002
|
| [17] |
Chaudary P, Marshall TI, Perozziello FM, et al. (2014) Relative biological effectiveness variation along monoenergetic and modulated bragg peaks of a 62-MeV therapeutic proton beam: a preclinical assessment. Int J Radiat Oncol Biol Phys 90: 27–35. doi: 10.1016/j.ijrobp.2014.05.010
|
| [18] |
Michaelidesová A, Vachelová J, Puchalska M, et al. (2017) Relative biological effectiveness in a proton spread-out Bragg peak formed by pencil beam scanning mode. Australas Phys Eng Sci Med 40: 359–368. doi: 10.1007/s13246-017-0540-8
|
| [19] |
Grün R, Friedrich T, Krämer M, et al. (2017) Systematics of relative biological effectiveness measurements for proton radiation along the spread out Bragg peak: experimental validation of the local effect model. Phys Med Biol 62: 890–908. doi: 10.1088/1361-6560/62/3/890
|
| [20] | Scholz M, Kraft G (1996) Track structure and the calculation of biological effects of heavy charged particles. Adv Space Res 18: 5–14. |
| [21] | Hubert F, Fleury A, Bimbot R, et al. (1980) Range and stopping power tables for 2.5–100 MeV/nucleon heavy ions in solids. Ann Phys 5: 214–218. |
| [22] | Nortllcliff LC, Shilling RF (1970) Range and stopping power tables for heavy ions. Atom Data Nucl Data 7: 233–437. |
| [23] |
Elsässer T, Krämer M, Scholz M (2008) Accuracy of the local effect model for the prediction of biologic effects of carbon ion beams in vitro and in vivo. Int J Radiat Oncol Biol Phys 71: 866–872. doi: 10.1016/j.ijrobp.2008.02.037
|
| [24] |
Kiefer J, Straaten H (1986) A model of ion track structure based on classical collision dynamics. Phys Med Biol 31: 1201–1209. doi: 10.1088/0031-9155/31/11/002
|
| [25] |
Scholz M, Kellerer AM, Kraft-Weyrather W, et al. (1997) Computation of cell survival in heavy ion beams for therapy. The model and its approximation. Radiat Environ Bioph 36: 59–66. doi: 10.1007/s004110050055
|
| [26] |
Elsässer T, Scholz M (2007) Cluster effects within the local effect model. Radiat Res 167: 319–329. doi: 10.1667/RR0467.1
|
| [27] |
Elsässer T, Weyrather WK, Friedrich T, et al. (2010) Quantification of the relative biological effectiveness for ion beam radiotherapy: direct experimental comparison of proton and carbon ion beams and a novel approach for treatment planning. Int J Radiat Oncol Biol Phys 78: 1177–1183. doi: 10.1016/j.ijrobp.2010.05.014
|
| [28] |
Grün R, Friedrich T, Elsässer T, et al. (2012) Impact of enhancements in the Local Effect Model (LEM) on the predicted RBE-weighted target dose distribution in carbon ion therapy. Phys Med Biol 57: 7261–7274. doi: 10.1088/0031-9155/57/22/7261
|
| [29] |
Kanai T, Furusawa Y, Fukutsu K, et al. (1997) Irradiation of mixed beam and design of spread-out Bragg peak for heavy-ion radiotherapy. Radiat Res 147: 78–85. doi: 10.2307/3579446
|
| [30] |
Kanai T, Endo M, Minohara S, et al (1999) Biophysical characteristics of HIMAC clinical irradiation system for heavy-ion radiation therapy. Int J Radiat Oncol Biol Phys 44: 201–210. doi: 10.1016/S0360-3016(98)00544-6
|
| [31] | Kraft G (2002) Radiobiological effecs of highly-charged ions, In: Currel FJ, The physics of highly and multiply charged ions, Kluwer Academic Publisher. |
| [32] |
Mizoe J, Tsujii H, Kamada T, et al. (2004) Dose escalation study of carbon ion radiotherapy for locally advanced head-and-neck cancer. Int J Radiat Oncol Biol Phys 60: 358–364. doi: 10.1016/j.ijrobp.2004.02.067
|
| [33] |
Kanai T, Matsufuji N, Miyamoto T, et al. (2006) Examination of GyE system for HIMAC carbon therapy. Int J Radiat Oncol Biol Phys 64: 650–656. doi: 10.1016/j.ijrobp.2005.09.043
|
| [34] |
Hawkins RB (1994) A statistical theory of cell killing by radiation of varying linear energy transfer. Radiat Res 140: 366–374. doi: 10.2307/3579114
|
| [35] |
Hawkins RB (2003) A microdosimetric-kinetic model for the effect of non-Poisson distribution of lethal lesions on the variation of RBE with LET. Radiat Res 160: 61–69. doi: 10.1667/RR3010
|
| [36] |
Kellerer M, Rossi HH (1978) A generalized formulation of dual radiation action. Radiat Res 75: 471–488. doi: 10.2307/3574835
|
| [37] |
Kase Y, Kanai T, Matsufuji N, et al. (2008) Biophysical calculation of cell survival probabilities using amorphous track structure models for heavy-ion irradiation. Phys Med Biol 53: 37–59. doi: 10.1088/0031-9155/53/1/003
|
| [38] |
Inaniwa T, Furukawa T, Kase Y, et al. (2010) Treatment planning for a scanned carbon beam with a modified microdosimetric kinetic model. Phys Med Biol 55: 6721–6737. doi: 10.1088/0031-9155/55/22/008
|
| [39] |
Furusawa Y, Fukutsu K, Aoki M, et al. (2000) Inactivation of aerobic and hypoxic cells from three different cell lines by accelerated He-, C- and Ne-ion beams. Radiat Res 154: 485–496. doi: 10.1667/0033-7587(2000)154[0485:IOAAHC]2.0.CO;2
|
| [40] |
Inaniwa T, Furukawa T, Sato S, et al. (2008) Development of treatment planning for scanning irradiation at HIMAC. Nucl Instr Meth Phys Res B 266: 2194–2198. doi: 10.1016/j.nimb.2008.02.070
|
| [41] |
Sato T, Kase Y, Watanabe R, et al. (2009) Biological dose estimation for charged particle therapy using an improved PHITS code coupled with a MK model. Radiat Res 171: 107–117. doi: 10.1667/RR1510.1
|
| [42] |
Sato T, Furusawa Y (2012) Cell survival fraction estimation based on the probability densities of domain and cell nucleus specific energies using improved Microdosimetric Kinetic Models. Radiat Res 178: 341–356. doi: 10.1667/RR2842.1
|
| [43] |
Sato T, Hamada N (2014) Model assembly for estimating cell surviving fraction for both targeted and nontargeted effects based on microdosimetric probability densities. P loS One 9: e114056. doi: 10.1371/journal.pone.0114056
|
| [44] |
Kramer M, Wang JF, Weyrather W (2003) Biological dosimetry of complex ion radiation fields. Phys Med Biol 48: 2063–2070. doi: 10.1088/0031-9155/48/14/302
|
| [45] |
Magro G, Dahle TJ, Molinelli S, et al. (2017) The FLUKA Monte Carlo code coupled with the NIRS approach for clinical dose calculations in carbon ion therapy. Phys Med Biol 62: 3814–3827. doi: 10.1088/1361-6560/aa642b
|
| [46] |
Frese MC, Yu VK, Stewart RD, et al. (2012) A mechanism-based approach to predict the relative biological effectiveness of protons and carbon ions in radiation therapy. Int J Radiat Oncol Biol Phys 83: 442–450. doi: 10.1016/j.ijrobp.2011.06.1983
|
| [47] |
Tilly N, Johansson J, Isacsson U, et al. (2005) The influence of RBE variations in clinical proton treatment plan for a hypopharynx cancer. Phys Med Biol 50: 2765–2777. doi: 10.1088/0031-9155/50/12/003
|
| [48] |
Belli M, Cera F, Cherubini R, et al. (1993) Inactivation and mutation induction in V79 cells by low energy protons: re-evaluation of the results at the LNL facility. Int J Radiat Biol 63: 331–337. doi: 10.1080/09553009314550441
|
| [49] |
Belli M, Cera F, Cherubini R, et al. (1998) RBE-LET relationships for cell inactivation and mutation induced by low energy protons in V79 cells: further results at the LNL facility. Int J Radiat Biol 74: 501–509. doi: 10.1080/095530098141375
|
| [50] |
Belli M, Bettega D, Calzolari P, et al. (2000) Inactivation of human normal and tumour cells irradiated with low energy protons. Int J Radiat Biol 76: 831–839. doi: 10.1080/09553000050028995
|
| [51] |
Folkard M, Prise KM, Vojnovic B, et al. (1996) Inactivation of V79 cells by low-energy protons, deuterons and helium-3 ions. Int J Radiat Biol 69: 729–738. doi: 10.1080/095530096145472
|
| [52] |
Bettega D, Calzolari P, Marchesini R, et al. (1998) Inactivation of C3H10T1/2 cells by low energy protons and deuterons. Int J Radiat Biol 73: 303–309. doi: 10.1080/095530098142400
|
| [53] | Frese MC, Wilkens JJ, Huber PE, et al. (2011) Application of constant vs. variable relative biological effectiveness in treatment planning of intensity-modulated proton therapy. Int J Radiat Oncol Biol Phys 79: 80–88. |
| [54] |
Wilkens JJ, Oelfke U (2004) A phenomenological model for the relative biological effectiveness in therapeutic proton beams. Phys Med Biol 49: 2811–2825. doi: 10.1088/0031-9155/49/13/004
|
| [55] | ICRU (1993) Stopping powers and ranges for protons and alpha particles, report 49, International Commission on Radiation Units and Measurements, Bethesda, MD, USA. |
| [56] |
Carabe-Fernandez A, Moteabbed M, Depauw N, et al. (2012) Range uncertainty in proton therapy due to variable biological effectiveness. Phys Med Biol 57: 1159–1172. doi: 10.1088/0031-9155/57/5/1159
|
| [57] |
Belli M, Cherubini R, Finotto S, et al. (1989) RBE-LET relationship for the survival of V79 cells irradiated with low energy protons. Int J Radiat Biol 55: 93–104. doi: 10.1080/09553008914550101
|
| [58] | Perris A, Pialoglou P, Katsanos AA, et al. (1986) Biological effectiveness of low energy protons. I. Survival of chinese hamster cells. Int J Radiat Biol 50: 1093–1101. |
| [59] |
Coutrakon G, Cortese J, Ghebremedhin A, et al. (1997) Microdosimetry spectra of the Loma Linda proton beam and relative biological effectiveness comparisons. Med Phys 24: 1499–1506. doi: 10.1118/1.598038
|
| [60] |
Wedenberg M, Lind BK, Hardemark B (2013) A model for the relative biological effectiveness of protons: the tissue specific parameter a/b of photons is a predictor for the sensitivity to LET changes. Acta Oncol 52: 580–588. doi: 10.3109/0284186X.2012.705892
|
| [61] | Paganetti H (2014) Relative biological effectiveness (RBE) values for proton beam therapy. Variations as a function of biological endpoint, dose, and linear energy transfer. Phys Med Biol 59: R419–R472. |
| [62] |
McNamara AL, Schuemann J, Paganetti H (2015) A phenomenological relative biological effectiveness (RBE) model for proton therapy based on all published in vitro cell survival data. Phys Med Biol 60: 8399–8416. doi: 10.1088/0031-9155/60/21/8399
|
| [63] |
Wedenberg M, Toma-Dasu I (2014) Disregarding RBE variation in treatment plan comparison may lead to bias in favor of proton plans. Med Phys 41: 091706. doi: 10.1118/1.4892930
|
| [64] |
Giovannini G, Böhlen T, Cabal G, et al. (2016) Variable RBE in proton therapy: comparison of different model predictions and their influence on clinical-like scenarios. Radiat Oncol 11: 68. doi: 10.1186/s13014-016-0642-6
|
| [65] |
Ottolenghi A, Ballarini F, Biaggi M (2001) Modelling chromosomal aberration induction by ionising radiation: the influence of interphase chromosome architecture. Adv Space Res 27: 369–382. doi: 10.1016/S0273-1177(01)00004-7
|
| [66] |
Ballarini F, Ottolenghi A (2004) A model of chromosome aberration induction and CML incidence at low doses. Radiat Environ Bioph 43: 165–171. doi: 10.1007/s00411-004-0246-7
|
| [67] | Ballarini F (2010) From DNA radiation damage to cell death: Theoretical approaches. J Nucleic Acids. |
| [68] |
Ballarini F, Altieri S, Bortolussi S, et al. (2013) A model of radiation-induced cell killing: insights into mechanisms and applications for hadrontherapy. Radiat Res 180: 307–315. doi: 10.1667/RR3285.1
|
| [69] |
Ballarini F, Altieri S, Bortolussi S, et al. (2014) The BIANCA model/code of radiation-induced cell death: application to human cells exposed to different radiation types. Radiat Environ Bioph 53: 525–533. doi: 10.1007/s00411-014-0537-6
|
| [70] |
Carante MP, Altieri S, Bortolussi S, at al. (2015) Modelling radiation-induced cell death: role of different levels of DNA damage clustering. Radiat Environ Bioph 54: 305–316. doi: 10.1007/s00411-015-0601-x
|
| [71] | Carante MP, Ballarini F (2016) Calculating variations in biological effectiveness for a 62 MeV proton beam. Front Oncol 6: 76. |
| [72] | Ballarini F, Carante MP (2016) Chromosome aberrations and cell death by ionizing radiation: evolution of a biophysical model. Radiat Phys Chem 128C: 18–25. |
| [73] |
Aiginger HV, Andersen F, Ballarini, et al. (2005) The FLUKA code: new developments and application to 1 GeV/n Iron beams. Adv Space Res 35: 214–222. doi: 10.1016/j.asr.2005.01.090
|
| [74] |
Ballarini F, Ottolenghi A (2005) A model of chromosome aberration induction: applications to space research. Radiat Res 164: 567–570. doi: 10.1667/RR3365.1
|
| [75] | Ballarini F, Alloni D, Facoetti A, et al. (2007) Radiation risk estimation: modelling approaches for "targeted" and "non-targeted" effects. Adv Space Res 40: 1392–1400. |
| [76] |
Campa AD, Alloni F, Antonelli, et al. (2009) DNA fragmentation induced in human fibroblasts by 56Fe ions: experimental data and MC simulations. Radiat Res 171: 438–445. doi: 10.1667/RR1442.1
|
| [77] |
Wu H, George K, Yang TC (1999) Estimate of the frequency of true incomplete exchanges in human lymphocytes exposed to 1 GeV/u Fe ions in vitro. Int J Radiat Biol 75: 593–599. doi: 10.1080/095530099140230
|
| [78] |
Cornforth MN, Bedford JS (1985) On the nature of a defect in cells from individuals with Ataxia-Telangiectasia. Science 227: 1589–1591. doi: 10.1126/science.3975628
|
| [79] |
Boei JJ, Vermeulen S, Mullenders LH, et al. (2001) Impact of radiation quality on the spectrum of induced chromosome exchange aberrations. Int J Radiat Biol 77: 847–857. doi: 10.1080/09553000110054610
|
| [80] |
Chen AM, Lucas JN, Simpson PJS, et al. (1997) Computer simulation of data on chromosome aberrations produced by X rays or alpha particles and detected by fluorescence in situ hybridization. Radiat Res 148: S93–S101. doi: 10.2307/3579722
|
| [81] |
Cornforth M, Bedford J (1987) A quantitative comparison of potentially lethal damage repair and the rejoining of interphase chromosome breaks in low passage normal human fibroblasts. Radiat Res 111: 385–405. doi: 10.2307/3576926
|
| [82] | Carrano AV (1973) Chromosome aberrations and radiation-induced cell death. I. Transmission and survival parameters of aberrations. Mutat Res 17: 341–353. |
| [83] |
Cuttone G, Cirrone GAP, Di FG, et al. (2011) CATANA protontherapy facility: The state of art of clinical and dosimetric experience. Eur Phys J Plus 126: 65. doi: 10.1140/epjp/i2011-11065-1
|
| [84] |
Rabbits T (1994) Chromosomal translocations in human cancer. Nature 372: 143–149. doi: 10.1038/372143a0
|
| [85] |
Tommasino F, Durante M (2015) Proton radiobiology. Cancers 7: 353–381. doi: 10.3390/cancers7010353
|
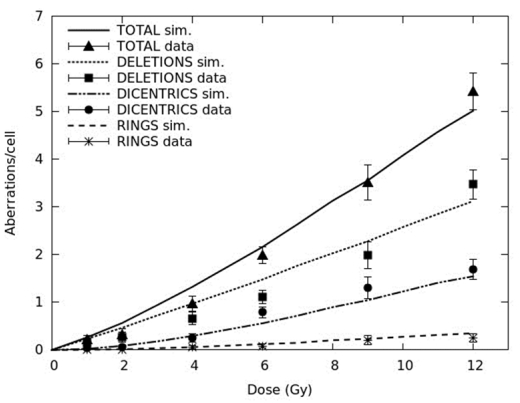
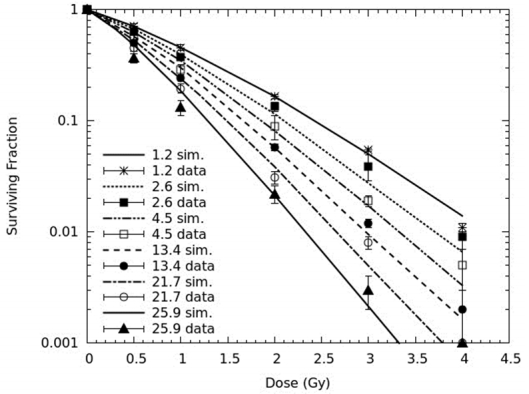
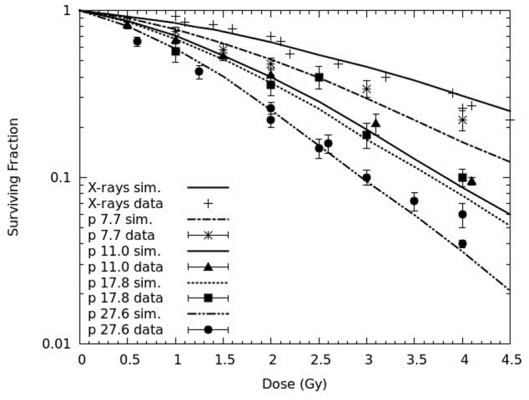
Figures(3) / Tables(4)

Mario P. Carante, Francesca Ballarini. Modelling cell death for cancer hadrontherapy[J]. AIMS Biophysics, 2017, 4(3): 465-490. doi: 10.3934/biophy.2017.3.465







 DownLoad:
DownLoad: