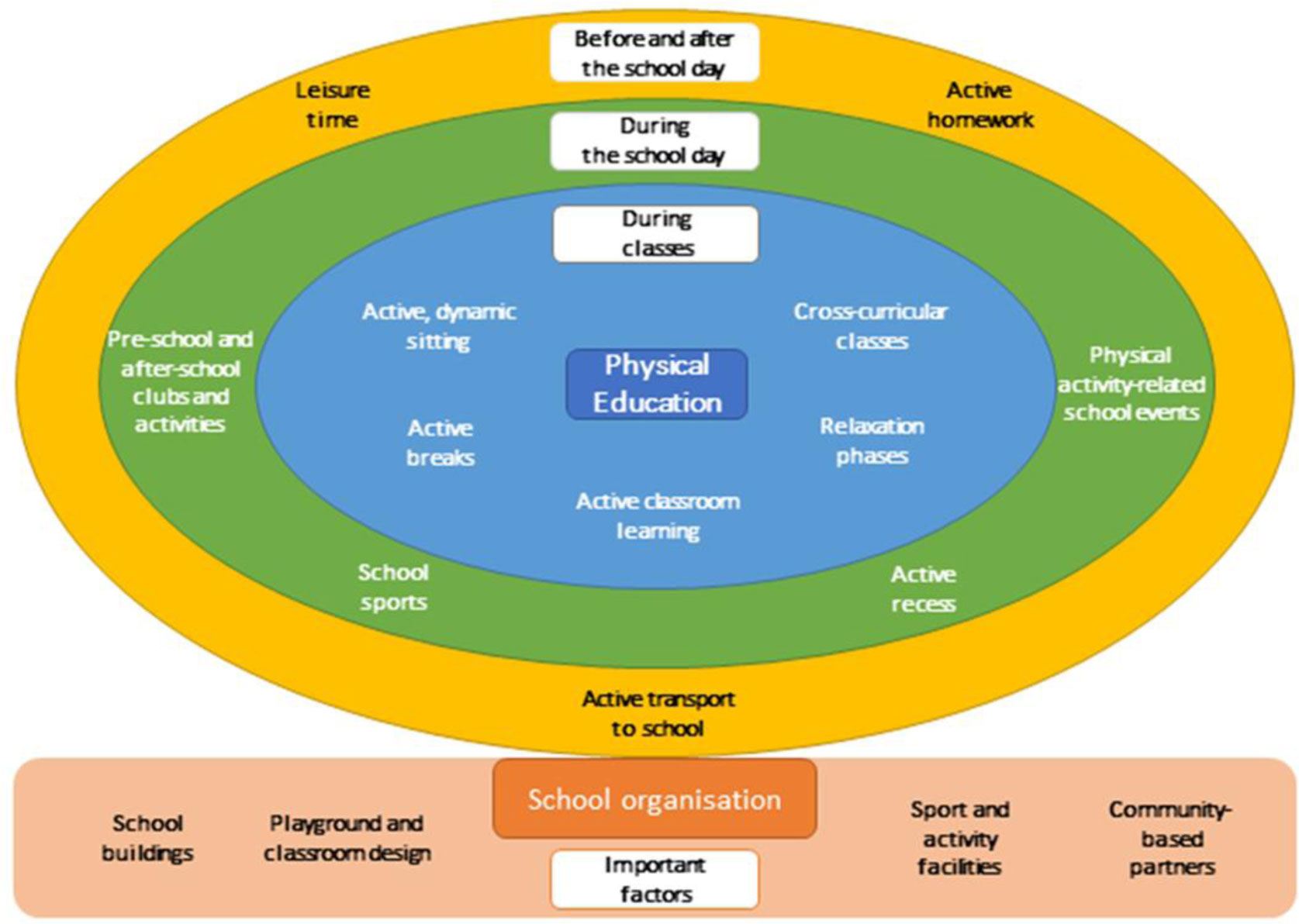
Measures devised to contain the COVID-19, including isolation, social distancing, and quarantine, have profoundly affected people's lives around the world. One of the consequences of these actions has been a general reduction in the habitual daily physical activity among children and young people for whom schools represent the major setting for the promotion of sports, physically active play, movement skills learning, and other activity supportive of healthy, active lifestyles. Whilst acknowledging the seriousness of these changes, and their concomitant health risks, we suggest that COVID-19 offers an opportunity to think again about important features of school-based activity promotion in light of new lessons learnt during lockdown, emerging technologies, and adapted pedagogies. In these specific cases, COVID-19 could be judged a “fortuitous disruptor” to the extent that it has opened a window of opportunity to schools and teachers to reflect on their assumptions about the scope, content, and delivery of their curricula, and on the new professional knowledge that has emerged. Active Homework, or physical activity-related tasks assigned to students by teachers that are meant to be carried out before, after and away from school, that students can do on their own or with family members, is not a new idea, but the enforced changes to school provision have made it considerably more common since the pandemic. Perhaps Active Homework is a concept worth retaining as schools start to return to “normal”? We offer a typology of Active Homework, and examine opportunities to expand, extend, and enhance physical education and physical activity opportunities by breaking down the presumed boundary between school and home. In conclusion, we suggest that Active Homework is worth exploring as a potentially valuable approach to enhancing the quantity and quality of students' school-based health-related physical activity. If so, considerably more research and curriculum development is needed.
Citation: Richard Bailey, Claude Scheuer. The COVID-19 pandemic as a fortuitous disruptor in physical education: the case of active homework[J]. AIMS Public Health, 2022, 9(2): 423-439. doi: 10.3934/publichealth.2022029
Measures devised to contain the COVID-19, including isolation, social distancing, and quarantine, have profoundly affected people's lives around the world. One of the consequences of these actions has been a general reduction in the habitual daily physical activity among children and young people for whom schools represent the major setting for the promotion of sports, physically active play, movement skills learning, and other activity supportive of healthy, active lifestyles. Whilst acknowledging the seriousness of these changes, and their concomitant health risks, we suggest that COVID-19 offers an opportunity to think again about important features of school-based activity promotion in light of new lessons learnt during lockdown, emerging technologies, and adapted pedagogies. In these specific cases, COVID-19 could be judged a “fortuitous disruptor” to the extent that it has opened a window of opportunity to schools and teachers to reflect on their assumptions about the scope, content, and delivery of their curricula, and on the new professional knowledge that has emerged. Active Homework, or physical activity-related tasks assigned to students by teachers that are meant to be carried out before, after and away from school, that students can do on their own or with family members, is not a new idea, but the enforced changes to school provision have made it considerably more common since the pandemic. Perhaps Active Homework is a concept worth retaining as schools start to return to “normal”? We offer a typology of Active Homework, and examine opportunities to expand, extend, and enhance physical education and physical activity opportunities by breaking down the presumed boundary between school and home. In conclusion, we suggest that Active Homework is worth exploring as a potentially valuable approach to enhancing the quantity and quality of students' school-based health-related physical activity. If so, considerably more research and curriculum development is needed.
Active homework
Health enhancing physical activity
Physical education

| [1] | Lincoln A Annual Message to Congress, Washington, DC: December 1, 1862 (1862). Available from: http://www.https://hd.housedivided.dickinson.edu/node/40533 |
| [2] | World Bank (2022) Global economic prospects. Washington, DC: World Bank Group. |
| [3] | Meyerowitz-Katz G, Bhatt S, Ratmann O, et al. (2021) Is the cure really worse than the disease? The health impacts of lockdowns during COVID-19. BMJ Glob Health 6: e006653. https://doi.org/10.1136/bmjgh-2021-006653 |
| [4] | Pietrobelli A, Pecoraro L, Ferruzzi A, et al. (2020) Effects of COVID-19 lockdown on lifestyle behaviors in children with obesity living in Verona, Italy: a longitudinal study. Obesity 28: 1382-1385. https://doi.org/10.1002/oby.22861 |
| [5] | Ravens-Sieberer U, Kaman A, Erhart M, et al. (2021) Impact of the COVID-19 pandemic on quality of life and mental health in children and adolescents in Germany. Eur Child Adoles Psy 1–11. https//doi.org/10.1007/s00787-021-01726-5 |
| [6] | Schwab K (2016) The fourth industrial revolution. Geneva: World Economic Forum. |
| [7] | UNICEFEducation disrupted: The second year of the COVID-19 pandemic and school closures (2021). Available from: https://data.unicef.org/resources/education-disrupted |
| [8] | Goldfeld S, O'Connor E, Sung V, et al. (2022) Potential indirect impacts of the COVID-19 pandemic on children: a narrative review using a community child health lens. Med J Aust 216: 364-372. https://doi.org/10.5694/mja2.51368 |
| [9] | Viner RM, Ward JL, Hudson LD, et al. (2021) Systematic review of reviews of symptoms and signs of COVID-19 in children and adolescents. Arch Dis Child 106: 802-807. https://doi.org/10.1136/archdischild-2020-320972 |
| [10] | OECD (2021) La situation de l’enseignement scolaire, Paris: OECD. |
| [11] | World Bank, UNESCO and UNICEFThe state of the global education crisis: A path to recovery (2021). Available from: https://www.unicef.org/reports/state-global-education-crisis |
| [12] | Bloom BS (1956) Taxonomy of educational objectives: The classification of educational goals. New York: Longmans Group. |
| [13] | (2015) UNESCOInternational Charter of Physical Education, Physical Activity and Sport. Paris: UNESCO. |
| [14] | Kristjánsson K (2019) Flourishing as the aim of education: A neo-Aristotelian view. London: Routledge. https://doi.org/10.4324/9780429464898 |
| [15] | Dunton GF, Do B, Wang SD (2020) Early effects of the COVID-19 pandemic on physical activity and sedentary behavior in children living in the US. BMC Public Health 20: 1351. https://doi.org/10.1186/s12889-020-09429-3 |
| [16] | Schmidt SCE, Burchatz A, Kolb S, et al. (2021) Zur Situation der körperlich-sportlichen Aktivität von Kindern und Jugendlichen während der COVID-19 Pandemie in Deutschland: Die Motorik-Modul Studie (MoMo). Karlsruher Institut für Technologie (KIT) Scientific Working Papers. |
| [17] | Wunsch K, Nigg C, Niessner C, et al. (2021) The impact of COVID-19 on the interrelation of physical activity, screen time and health-related quality of Life in children and adolescents in Germany: results of Motorik-Modul Study. Children 8: 98. https://doi.org/10.3390/children8020098 |
| [18] | Derigny T, Schnitzler C, Gandrieau J, et al. (2022) Resilience of adolescents in physical activity during the covid-19 pandemic: a preliminary case study in France. Phys Act Rev 10: 86-97. https://doi.org/10.16926/rp.2020.12.10 |
| [19] | Zenic N, Taiar R, Gilic B, et al. (2020) Levels and changes of physical activity in adolescents during the COVID-19 Pandemic: Contextualizing urban vs. rural living environment. Appl Sci 10: 3997. https://doi.org/10.3390/app10113997 |
| [20] | Margaritis I, Houdart S, El Ouadrhiri Y, et al. (2020) How to deal with COVID-19 epidemic-related lockdown physical inactivity and sedentary increase in youth?. Arch Public Health 78: 52. https://doi.org/10.1186/s13690-020-00432-z |
| [21] | Vanderloo LM, Carsley S, Aglipay M, et al. (2020) Applying harm reduction principles to address screen time in young children amidst the COVID-19 pandemic. J Dev Behav Pediatr 41: 335-336. https://doi.org/10.1097/DBP.0000000000000825 |
| [22] | Moore SA, Faulkner G, Rhodes RE, et al. (2020) Impact of the COVID-19 virus outbreak on movement and play behaviours of Canadian children and youth. Int J Behav Nutr Phys Act 17: 85. https://doi.org/10.1186/s12966-020-00987-8 |
| [23] | Xiang M, Zhang Z, Kuwahara K (2020) Impact of COVID-19 pandemic on children and adolescents' lifestyle behavior larger than expected. Prog Cardiovasc Dis 63: 531-532. https://doi.org/10.1016/j.pcad.2020.04.013 |
| [24] | Ten Velde G, Lubrecht J, Arayess L, et al. (2021) Physical activity behaviour and screen time in Dutch children during the COVID-19 pandemic: Pre-, during-and post-school closures. Pediatric Obes 16: e12779. https://doi.org/10.1111/ijpo.12779 |
| [25] | Greier K, Drenowatz C, Bischofer T, et al. (2021) Physical activity and sitting time prior to and during COVID-19 lockdown in Austrian high-school students. AIMS Public Health 8: 531. https://doi.org/10.3934/publichealth.2021043 |
| [26] | Hyunshik K, Jiameng M, Sunkyoung L, et al. (2021) Change in Japanese children's 24-hour movement guidelines and mental health during the COVID-19 pandemic. Sci Rep 11: 1-9. https://doi.org/10.1038/s41598-021-01803-4 |
| [27] | Ostermeier E, Tucker P, Clark A, et al. (2021) Parents' report of Canadian elementary school children's physical activity and screen time during the COVID-19 pandemic. Int J Environ Res Public Health 18: 12352. https://doi.org/10.3390/ijerph182312352 |
| [28] | Grgic J, Dumuid D, Bengoechea EG, et al. (2018) Health outcomes associated with reallocations of time between sleep, sedentary behaviour, and physical activity. Int J Behav Nutr Phys Act 15: 69. https://doi.org/10.1186/s12966-018-0691-3 |
| [29] | Breidokienė R, Jusienė R, Urbonas V, et al. (2021) Sedentary behavior among 6–14-Year-Old children during the COVID-19 lockdown and its relation to physical and mental health. Healthcare 9: 756. https://doi.org/10.3390/healthcare9060756 |
| [30] | Van Sluijs EM, Ekelund U, Crochemore-Silva I, et al. (2021) Physical activity behaviours in adolescence: current evidence and opportunities for intervention. Lancet 398: 429-442. https://doi.org/10.1016/S0140-6736(21)01259-9 |
| [31] | Okazaki K, Suzuki K, Sakamoto Y, et al. (2015) Physical activity and sedentary behavior among children and adolescents living in an area affected by the 2011 Great East Japan earthquake and tsunami for 3 years. Prev Med Rep 2: 720-724. https://doi.org/10.1016/j.pmedr.2015.08.010 |
| [32] | Cope E, Bailey RP (2016) A review of research and evidence on factors that impact on children's sport and physical activity behaviours and attitudes. London: Sport England. |
| [33] | Daly-Smith A, Quarmby T, Archbold VS, et al. (2020) Using a multi-stakeholder experience-based design process to co-develop the Creating Active Schools Framework. Int J Behav Nutr Phys Act 17: 1-12. https://doi.org/10.1186/s12966-020-0917-z |
| [34] | Bailey RP (2018) Is sport good for us?. Sport and Health . London: Routledge 7-35. https://doi.org/10.4324/9781315266459-2 |
| [35] | McKenzie TL, Lounsbery MA (2014) The pill not taken: Revisiting physical education teacher effectiveness in a public health context. Res Q Exerc Sport 85: 287-292. https://doi.org/10.1080/02701367.2014.931203 |
| [36] | Bailey RP, Vašíčková J, Vlček P, et al. (2019) An international review of the contributions of school-based physical activity, physical education, and school sport to the promotion of health-enhancing physical activity. Timeline . https://doi.org/10.5281/zenodo.5899571 |
| [37] | Gobbi E, Maltagliati S, Sarrazin P, et al. (2020) Promoting physical activity during school closures imposed by the first wave of the COVID-19 pandemic. Int J Environ Res Public Health 17: 9431. https://doi.org/10.3390/ijerph17249431 |
| [38] | Varea V, González-Calvo G, García-Monge A (2022) Exploring the changes of physical education in the age of Covid-19. Phys Educ Sport Pedagogy 27: 32-42. https://doi.org/10.1080/17408989.2020.1861233 |
| [39] | Ding D, Sallis JF, Kerr J, et al. (2011) Neighborhood environment and physical activity among youth: a review. Am J Prev Med 41: 442-455. https://doi.org/10.1016/j.amepre.2011.06.036 |
| [40] | Masonbrink AR, Hurley E (2020) Advocating for children during the COVID-19 school closures. Pediatrics 146: e20201440. https://doi.org/10.1542/peds.2020-1440 |
| [41] | Bailey R, Hillman C, Arent S, et al. (2013) Physical activity: an underestimated investment in human capital?. J Phys Act Health 10: 289-308. https://doi.org/10.1123/jpah.10.3.289 |
| [42] | Schmidt SC, Anedda B, Burchartz A, et al. (2020) Physical activity and screen time of children and adolescents before and during the COVID-19 lockdown in Germany. Sci Rep 10: 1-12. https://doi.org/10.1038/s41598-020-78438-4 |
| [43] | Ding D, del Pozo Cruz B, Green MA, et al. (2020) Is the COVID-19 lockdown nudging people to be more active. Br J Sports Med 54: 1183-1184. https://doi.org/10.1136/bjsports-2020-102575 |
| [44] | Schnaiderman D, Bailac M, Borak L, et al. (2021) Psychological impact of COVID-19 lockdown in children and adolescents from San Carlos de Bariloche, Argentina. Arch Argent Pediatr 119: 170-176. https://doi.org/10.5546/aap.2021.eng.170 |
| [45] | Perez D, Thalken JK, Ughelu NE, et al. (2021) Nowhere to go: Parents' descriptions of children's physical activity during a global pandemic. Front Public Health 9: 223. https://doi.org/10.3389/fpubh.2021.642932 |
| [46] | Davies BR, Wood L, Banfield K, et al. (2014) The provision of active after-school clubs for children in English primary schools: implications for increasing children's physical activity. Open J Prev Med . https://doi.org/10.4236/ojpm.2014.47069 |
| [47] | Elflein P, Huh Y (2014) Sport und Sportunterricht im Rahmen einer gesunden Schule. Die berufsbildende Sch 66: 168-173. |
| [48] | Maltagliati S, Carraro A, Escriva-Boulley G, et al. (2021) Changes in physical education teachers' motivations predict the evolution of behaviors promoting students' physical activity during the COVID-19 lockdown. SportRxiv . https://doi.org/10.31236/osf.io/h4mw8 |
| [49] | Zhu X, Liu J (2020) Education in and after Covid-19: Immediate responses and long-term visions. Postdigit Sci Educ 2: 695-699. https://doi.org/10.1007/s42438-020-00126-3 |
| [50] | Gadamer HG (1960) Wahrheit und Methode. Tübingen: Mohr. |
| [51] | Gard M, Pluim C (2014) Schools and public health: Past, present, future. Lexington Books. |
| [52] | Scheuer C, Bailey RP (2021) The active school concept. Physical Activity and Sport During the First Ten Years of Life . London: Routledge 173-187. https://doi.org/10.4324/9780429352645-19 |
| [53] | Illi U (1995) Bewegte Schule. Die Bedeutung Und Funktion Der Bewegung Als Beitrag Zu Einer Ganzheitlichen Gesundheitsbildung Im Lebensraum Schule. Sportunterricht 44: 404-415. |
| [54] | Thiel A, Teubert H (2018) Die Bewegte Schule. Handbuch Bildungsreform und Reformpädagogik . Wiesbaden: Springer Verlag. https://doi.org/10.1007/978-3-658-07491-3_47 |
| [55] | Fox KR (1996) Physical activity promotion and the active school. New directions in physical education . London: Farmer 94-109. |
| [56] | Fox K, Harris J (2003) Promoting physical activity through schools. Perspectives on health and exercise . Basingstoke: Palgrave Macmillan 181-201. |
| [57] | Bailey RP, Heck S, Scheuer C (2022) Active Homework: an under-researched element of the active schools concept. Ger J Exerc Sport Res 1–7. https://doi.org/10.1007/s12662-022-00802-5 |
| [58] | Smith MA, Claxton DB (2003) Using active homework in physical education. J Phys Educ Recreat 74: 28-32. https://doi.org/10.1080/07303084.2003.10608482 |
| [59] | St Ours E, Scrabis-Fletcher KA (2013) Implementing active homework in secondary physical education. Strategies 26: 23-27. https://doi.org/10.1080/08924562.2013.839433 |
| [60] | Smith J, Cluphf D, O'Connor J (2001) Homework in elementary physical education: a pilot study. Percept Mot Skills 92: 133-136. https://doi.org/10.2466/pms.2001.92.1.133 |
| [61] | Kohl HW, Cook HD (2012) Educating the student body: Taking physical activity and physical education to school. Washington DC: National Academies Press. |
| [62] | Storey KE, Montemurro G, Flynn J, et al. (2016) Essential conditions for the implementation of comprehensive school health to achieve changes in school culture and improvements in health behaviours of students. BMC Public Health 16: 1133. https://doi.org/10.1186/s12889-016-3787-1 |
| [63] | Beets MW, Okely A, Weaver RG, et al. (2016) The theory of expanded, extended, and enhanced opportunities for youth physical activity promotion. Int J Behav Nutr Phys Act 13: 1-15. https://doi.org/10.1186/s12966-016-0442-2 |
| [64] | Terhart E (2009) Didaktik. Stuttgart: Philipp Reclam. |
| [65] | Bull F, Al-Ansari SS, Biddle S, et al. (2020) World Health Organization 2020 guidelines on physical activity and sedentary behaviour. Br J Sports Med 54: 1451-1462. https://doi.org/10.1136/bjsports-2020-102955 |
| [66] | Wagner I (2016) Wissen im Sportunterricht. Aachen: Meyer & Meyer Verlag. https://doi.org/10.5771/9783840311796 |
| [67] | Kääpä M, Palomäki S, Vähä-Ypyä H, et al. (2019) The role of physical education homework to adolescent girls' physical activity in Finland. Adv Phys Educ 9: 223-239. https://doi.org/10.4236/ape.2019.94016 |
| [68] | Cooper H (1989) Homework. White Plains, NY: Longman. https://doi.org/10.1037/11578-000 |
| [69] | Cooper H, Robinson JC, Patall EA (2006) Does homework improve academic achievement? A synthesis of research. Rev Educ Res 76: 1-62. https://doi.org/10.3102/00346543076001001 |
| [70] | Vogel P (2007) Hausaufgaben in einer Bewegten Schule. Bewegte Schule: Schule bewegt gestalten . Baltmannsweiler: Schneider-Verlag Hohengehren 246-259. |
| [71] | (2011) BASPO (Bundesamt für Sport (BASPO)Die bewegte Schule. Erklärungen zum Schweizer Modell . Magglingen: BASPO. |
| [72] | Duncan S, Stewart T, McPhee J, et al. (2019) Efficacy of a compulsory homework programme for increasing physical activity and improving nutrition in children: a cluster randomised controlled trial. Int J Behav Nutr Phys Act 16: 80. https://doi.org/10.1186/s12966-019-0840-3 |
| [73] | Williams SM, Hannon JC (2013) Physical education homework that involves the family. Strategies 26: 3-8. https://doi.org/10.1080/08924562.2013.779848 |
| [74] | Brown TD, Penney D (2017) Examination physical education: policy, practice and possibilities. London: Routledge. https://doi.org/10.4324/9781315753997 |
| [75] | Hill K (2018) Homework in physical education?. J Phys Educ Recreat Dance 89: 58-63. https://doi.org/10.1080/07303084.2018.1440263 |
| [76] | Kinchin GD, O'Sullivan M (2003) Incidences of student support for and resistance to a curricular innovation in high school physical education. J Teach Phys Educ 22: 245-260. https://doi.org/10.1123/jtpe.22.3.245 |
| [77] | Smith M, Madden M (2014) Middle School Students' Reactions to the Implementation of Active Homework in Physical Education. Global J Health Phys Educ Pedagog . |
| [78] | Smith J, Cluphf D, O'Connor J (2001) Homework in elementary physical education. Percept Mot Skills 92: 133-136. https://doi.org/10.2466/pms.2001.92.1.133 |
| [79] | Pantanowitz M, Lidor R, Nemet D, et al. (2011) Feasibility of physical education homework assignments in high-school. Children and Exercise XXVII . London: Routledge 169-173. |
| [80] | Brazendale K, Beets MW, Weaver RG, et al. (2017) Understanding differences between summer vs. school obesogenic behaviors of children: the structured days hypothesis. Int J Behav Nutr Phys Act 14: 1-14. https://doi.org/10.1186/s12966-017-0555-2 |
| [81] | Moreno JP, Johnston CA, Woehler D (2013) Changes in weight over the school year and summer vacation. J Sch Health 83: 473-477. https://doi.org/10.1111/josh.12054 |
| [82] | Fairclough SJ, Boddy LM, Mackintosh KA, et al. (2015) Weekday and weekend sedentary time and physical activity in differentially active children. J Sci Med Sport 18: 444-449. https://doi.org/10.1016/j.jsams.2014.06.005 |
| [83] | Sailer M, Murböck J, Fischer F (2021) Digital learning in schools: What does it take beyond digital technology?. Teach Teach Educ 103: 103346. https://doi.org/10.1016/j.tate.2021.103346 |
| [84] | OECDDigital strategies in education across OECD countries: Exploring education policies on digital technologies (2020). Avaliable from: https://www.oecd.org/education/digital-strategies-in-education-across-oecd-countries-33dd4c26-en.htm |
| [85] | Scully D, Lehane P, Scully C (2021) ‘It is no longer scary’: digital learning before and during the Covid-19 pandemic in Irish secondary schools. Technol Pedagog Educ 30: 159-181. https://doi.org/10.1080/1475939X.2020.1854844 |
| [86] | Jie ND (2016) Towards a framework of education policy analysis. THF Literature Rev . https://doi.org/10.4324/9781003002666-8 |
| [87] | Bailey RP, Callary B (2022) Sport education from a global perspective. Education in Sport and Physical Activity: Future Directions and Global Perspectives . London: Routledge. |
| [88] | Kirk D (2009) Physical education futures. London: Routledge. https://doi.org/10.4324/9780203874622 |
| [89] | Colao A, Piscitelli P, Pulimeno M, et al. (2020) Rethinking the role of the school after COVID-19. Lancet Public Health 5: e370. https://doi.org/10.1016/S2468-2667(20)30124-9 |

Figures(2) / Tables(1)

Richard Bailey, Claude Scheuer. The COVID-19 pandemic as a fortuitous disruptor in physical education: the case of active homework[J]. AIMS Public Health, 2022, 9(2): 423-439. doi: 10.3934/publichealth.2022029







 DownLoad:
DownLoad: