Diabetes mellitus (DM) has a high incidence of comorbidities among patients with severe coronavirus disease 2019 (COVID-19). The elevated prevalence of DM in the world population makes it a significant risk factor because diabetic individuals appear to be prone to clinical complications and have increased mortality rates. Here, we review the possible underlying mechanisms involved in DM that led to worse outcomes in COVID-19. The impacts of hyperglycemia side effects, secondary comorbidities, weakened innate and adaptive immunity, chronic inflammation, and poor nutritional status, commonly present in DM, are discussed. The role of the SARS-CoV-2 receptor and its polymorphic variations on higher binding affinity to facilitate viral uptake in people with DM were also considered. Clinical differences between individuals with type 1 DM and type 2 DM affected by COVID-19 and the potential diabetogenic effect of SARS-CoV-2 infection were addressed.
Citation: María D Figueroa-Pizano, Alma C Campa-Mada, Elizabeth Carvajal-Millan, Karla G Martinez-Robinson, Agustin Rascon Chu. The underlying mechanisms for severe COVID-19 progression in people with diabetes mellitus: a critical review[J]. AIMS Public Health, 2021, 8(4): 720-742. doi: 10.3934/publichealth.2021057
Diabetes mellitus (DM) has a high incidence of comorbidities among patients with severe coronavirus disease 2019 (COVID-19). The elevated prevalence of DM in the world population makes it a significant risk factor because diabetic individuals appear to be prone to clinical complications and have increased mortality rates. Here, we review the possible underlying mechanisms involved in DM that led to worse outcomes in COVID-19. The impacts of hyperglycemia side effects, secondary comorbidities, weakened innate and adaptive immunity, chronic inflammation, and poor nutritional status, commonly present in DM, are discussed. The role of the SARS-CoV-2 receptor and its polymorphic variations on higher binding affinity to facilitate viral uptake in people with DM were also considered. Clinical differences between individuals with type 1 DM and type 2 DM affected by COVID-19 and the potential diabetogenic effect of SARS-CoV-2 infection were addressed.
Diabetes mellitus
Coronavirus disease 2019
Severe Acute Respiratory Syndrome Coronavirus
Severe Acute Respiratory Syndrome Coronavirus 2
Research Center for Food and Development
World Health Organization
Center for Systems Science and Engineering
Middle East respiratory syndrome
Intensive care unit
Angiotensin-converting enzyme 1
Angiotensin-converting enzyme 2
ACE inhibitors
Renin-angiotensin-aldosterone system
Angiotensin-receptor blockers
glycosylated hemoglobin
Reactive oxygen species
Cluster of differentiation 4+
Cluster of differentiation 8+
Tumor necrosis factor-α
Interleukin
C-reactive protein
Interferon-ϒ
Chest computed tomography
Lactate dehydrogenase levels
Receptor-binding domain
Ribonucleic acid
Desoxyribonucleic acid
Single nucleotide polymorphisms
Coronary heart disease
Body mass index
Diabetic ketoacidosis
transmembrane serine protease 2

| [1] |
Asselah T, Durantel D, Pasmant E, et al. (2021) COVID-19: discovery, diagnostics and drug development. J Hepatol 74: 168-184. doi: 10.1016/j.jhep.2020.09.031

|
| [2] |
Rojas R, Basto A, Aguilar C, et al. (2018) Prevalencia de diabetes por diagnóstico médico previo en México. Salud Publica Mex 60: 224-232. doi: 10.21149/8566
|
| [3] |
Levaillant M, Lièvre G, Baert G (2019) Ending diabetes in Mexico. Lancet 394: 467-468. doi: 10.1016/S0140-6736(19)31662-9
|
| [4] |
Singer M (2020) Deadly companions: COVID-19 and diabetes in Mexico. Med Anthropol 39: 660-665. doi: 10.1080/01459740.2020.1805742
|
| [5] |
Zhang W, Davis BD, Chen SS, et al. (2021) Emergence of a novel SARS-CoV-2 variant in southern California. JAMA 325: 1324-1326. doi: 10.1001/jama.2021.1612
|
| [6] |
Zucman N, Uhel F, Descamps D, et al. (2021) Severe reinfection with South African severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) variant 501Y.V2: a case report. Clin Infect Dis ciab129. doi: 10.1093/cid/ciab129
|
| [7] |
Fujino T, Nomoto H, Kutsuna S, et al. (2021) Novel SARS-CoV-2 variant in travelers from Brazil to Japan. Emerg Infect Dis 27: 1243-1245. doi: 10.3201/eid2704.210138
|
| [8] |
Yadav PD, Nyayanit DA, Sahay RR, et al. (2021) Isolation and characterization of the new SARS-CoV-2 variant in travellers from the United Kingdom to India: VUI-202012/01 of the B.1.1.7 lineage. J Travel Med 28: taab009. doi: 10.1093/jtm/taab009
|
| [9] |
Zuckerman NS, Fleishon S, Bucris E, et al. (2021) A unique SARS-CoV-2 spike protein P681H variant detected in Israel. Vaccines 9: 616. doi: 10.3390/vaccines9060616
|
| [10] | Munitz A, Yechezkel M, Dickstein Y, et al. (2021) The rise of SARS-CoV-2 variant B.1.1.7 in Israel intensifies the role of surveillance and vaccination in elderly. medRxiv 2021. |
| [11] | Mor O, Zuckerman NS, Hazan I, et al. (2021) BNT162b2 vaccination efficacy is marginally affected by the SARS-CoV-2 B.1.351 variant in fully vaccinated individuals. Avaliable at SSRN 3878825 . |
| [12] |
Munitz A, Yechezkel M, Dickstein Y, et al. (2021) Report BNT162b2 vaccination effectively prevents the rapid populations in Israel ll BNT162b2 vaccination effectively prevents the rapid rise of SARS-CoV-2 variant B.1.1.7 in high-risk populations in Israel. Cell Rep Med 2: 100264. doi: 10.1016/j.xcrm.2021.100264
|
| [13] |
Haas EJ, Angulo FJ, Mclaughlin JM, et al. (2021) Impact and effectiveness of mRNA BNT162b2 vaccine against SARS-CoV-2 infections and COVID-19 cases, hospitalisations, and deaths following a nationwide vaccination campaign in Israel: an observational study using national surveillance data. Lancet 397: 1819-1829. doi: 10.1016/S0140-6736(21)00947-8
|
| [14] |
Kustin T, Harel N, Finkel U, et al. (2021) Evidence for increased breakthrough rates of SARS-CoV-2 variants of concern in BNT162b2-mRNA-vaccinated individuals. Nat Med 27: 1379-1384. doi: 10.1038/s41591-021-01413-7
|
| [15] |
Hu T, Liu Y, Zhao M, et al. (2020) A comparison of COVID-19, SARS and MERS. PeerJ 8: e9725. doi: 10.7717/peerj.9725
|
| [16] |
Harrison AG, Lin T, Wang P (2020) Mechanisms of SARS-CoV-2 transmission and pathogenesis. Trends Immunol 41: 1100-1115. doi: 10.1016/j.it.2020.10.004
|
| [17] |
Yuki K, Fujiogi M, Koutsogiannaki S (2020) COVID-19 pathophysiology: a review. Clin Immunol 215: 108427. doi: 10.1016/j.clim.2020.108427
|
| [18] |
Sardu C, Gambardella J, Morelli MB, et al. (2020) Hypertension, thrombosis, kidney failure, and diabetes: is COVID-19 an endothelial disease? A comprehensive evaluation of clinical and basic evidence. J Clin Med 9: 1417. doi: 10.3390/jcm9051417
|
| [19] |
Safwat MA, Abdul-Rahman A, Forsan H, et al. (2021) COVID-19; Immunology, pathology, severity and immunosuppressants. Azhar Int J Pharm Med Sci 1: 1-14. doi: 10.21608/aijpms.2021.50103.1005
|
| [20] | Kabashneh S, Ali H, Alkassis S (2020) Multi-Organ failure in a patient with diabetes due to COVID-19 with clear lungs. Cureus 12: e8147. |
| [21] |
Guo L, Shi Z, Zhang Y, et al. (2020) Comorbid diabetes and the risk of disease severity or death among 8807 COVID-19 patients in China: a meta-analysis. Diabetes Res Clin Pract 166: 108346. doi: 10.1016/j.diabres.2020.108346
|
| [22] | Zhou Y, Chi J, Lv W, et al. (2021) Obesity and diabetes as high-risk factors for severe coronavirus disease 2019 (Covid-19). Diabetes Metab Res Rev 37: e3377. |
| [23] |
Parveen R, Sehar N, Bajpai R, et al. (2020) Association of diabetes and hypertension with disease severity in covid-19 patients: a systematic literature review and exploratory meta-analysis. Diabetes Res Clin Pract 166: 108295. doi: 10.1016/j.diabres.2020.108295
|
| [24] |
Frydrych LM, Bian G, O'Lone DE, et al. (2018) Obesity and type 2 diabetes mellitus drive immune dysfunction, infection development, and sepsis mortality. J Leukoc Biol 104: 525-534. doi: 10.1002/JLB.5VMR0118-021RR
|
| [25] |
Sur J, Sharma J, Sharma D (2021) Diabetes might augment the severity of COVID-19: a current prospects. Front Cardiovasc Med 7: 613255. doi: 10.3389/fcvm.2020.613255
|
| [26] |
Kumar A, Arora A, Sharma P, et al. (2020) Is diabetes mellitus associated with mortality and severity of COVID-19? A meta-analysis. Diabetes Metab Syndr 14: 535-545. doi: 10.1016/j.dsx.2020.04.044
|
| [27] |
Jeong IK, Yoon KH, Lee MK (2020) Diabetes and COVID-19: global and regional perspectives. Diabetes Res Clin Pract 166: 108303. doi: 10.1016/j.diabres.2020.108303
|
| [28] |
Chen Y, Yang D, Cheng B, et al. (2020) Clinical characteristics and outcomes of patients with diabetes and COVID-19 in association with glucose-lowering medication. Diabetes Care 43: 1399-1407. doi: 10.2337/dc20-0660
|
| [29] |
Mantovani A, Byrne CD, Zheng MH, et al. (2020) Diabetes as a risk factor for greater COVID-19 severity and in-hospital death: a meta-analysis of observational studies. Nutr Metab Cardiovasc Dis 30: 1236-1248. doi: 10.1016/j.numecd.2020.05.014
|
| [30] |
Du M, Lin YX, Yan WX, et al. (2020) Prevalence and impact of diabetes in patients with COVID-19 in China. World J Diabetes 11: 468-480. doi: 10.4239/wjd.v11.i10.468
|
| [31] |
Targher G, Mantovani A, Wang XB, et al. (2020) Patients with diabetes are at higher risk for severe illness from COVID-19. Diabetes Metab 46: 335-337. doi: 10.1016/j.diabet.2020.05.001
|
| [32] |
Ciardullo S, Zerbini F, Perra S, et al. (2020) Impact of diabetes on COVID-19-related in-hospital mortality: a retrospective study from Northern Italy. J Endocrinol Invest 44: 843-850. doi: 10.1007/s40618-020-01382-7
|
| [33] |
Halvatsiotis P, Kotanidou A, Tzannis K, et al. (2020) Demographic and clinical features of critically ill patients with COVID-19 in Greece: the burden of diabetes and obesity. Diabetes Res Clin Pract 166: 108331. doi: 10.1016/j.diabres.2020.108331
|
| [34] |
Nachtigall I, Lenga P, Jóźwiak K, et al. (2020) Clinical course and factors associated with outcomes among 1904 patients hospitalized with COVID-19 in Germany: an observational study. Clin Microbiol Infect 26: 1663-1669. doi: 10.1016/j.cmi.2020.08.011
|
| [35] |
Sutter W, Duceau B, Vignac M, et al. (2020) Association of diabetes and outcomes in patients with COVID-19: propensity score-matched analyses from a French retrospective cohort. Diabetes Metab 47: 101222. doi: 10.1016/j.diabet.2020.101222
|
| [36] |
Barron E, Bakhai C, Kar P, et al. (2020) Associations of type 1 and type 2 diabetes with COVID-19-related mortality in England: a whole-population study. Lancet Diabetes Endocrinol 8: 813-822. doi: 10.1016/S2213-8587(20)30272-2
|
| [37] |
Ge Y, Sun S, Shen Y (2021) Estimation of case-fatality rate in COVID-19 patients with hypertension and diabetes mellitus in the New York State: a preliminary report. Epidemiol Infect 149: e14. doi: 10.1017/S0950268821000066
|
| [38] |
Hussain S, Baxi H, Chand Jamali M, et al. (2020) Burden of diabetes mellitus and its impact on COVID-19 patients: a meta-analysis of real-world evidence. Diabetes Metab Syndr 14: 1595-1602. doi: 10.1016/j.dsx.2020.08.014
|
| [39] |
Ashktorab H, Pizuorno A, Oskroch G, et al. (2021) COVID-19 in Latin America: symptoms, morbidities, and gastrointestinal manifestations. Gastroenterology 160: 938-940. doi: 10.1053/j.gastro.2020.10.033
|
| [40] |
Izzi-Engbeaya C, Distaso W, Amin A, et al. (2021) Adverse outcomes in COVID-19 and diabetes: a retrospective cohort study from three London teaching hospitals. BMJ Open Diabetes Res Care 9: e001858. doi: 10.1136/bmjdrc-2020-001858
|
| [41] |
Ebekozien O, Agarwal S, Noor N, et al. (2020) Inequities in diabetic ketoacidosis among patients with type 1 diabetes and COVID-19: data from 52 US clinical centers. J Clin Endocrinol Metab 106: e1755-e1762. doi: 10.1210/clinem/dgaa920
|
| [42] |
Hafidh K, Abbas S, Khan A, et al. (2020) The clinical characteristics and outcomes of COVID-19 infections in patients with diabetes at a tertiary care center in the UAE. Dubai Diabetes Endocrinol J 26: 158-163. doi: 10.1159/000512232
|
| [43] |
Yan Y, Yang Y, Wang F, et al. (2020) Clinical characteristics and outcomes of patients with severe covid-19 with diabetes. BMJ Open Diabetes Res Care 8: e001343. doi: 10.1136/bmjdrc-2020-001343
|
| [44] |
Ling P, Luo S, Zheng X, et al. (2021) Elevated fasting blood glucose within the first week of hospitalization was associated with progression to severe illness of COVID-19 in patients with preexisting diabetes: a multicenter observational study. J Diabetes 13: 89-93. doi: 10.1111/1753-0407.13121
|
| [45] |
Silverio A, Di Maio M, Citro R, et al. (2021) Cardiovascular risk factors and mortality in hospitalized patients with COVID-19: systematic review and meta-analysis of 45 studies and 18,300 patients. BMC Cardiovasc Disord 21: 23. doi: 10.1186/s12872-020-01816-3
|
| [46] |
Acharya D, Lee K, Lee DS, et al. (2020) Mortality rate and predictors of mortality in hospitalized COVID-19 patients with diabetes. Healthcare 8: 338. doi: 10.3390/healthcare8030338
|
| [47] |
Bode B, Garrett V, Messler J, et al. (2020) Glycemic characteristics and clinical outcomes of COVID-19 patients hospitalized in the United States. J Diabetes Sci Technol 14: 813-821. doi: 10.1177/1932296820924469
|
| [48] | Bhandari S, Rankawat G, Singh A, et al. (2020) Impact of glycemic control in diabetes mellitus on management of COVID-19 infection. Int J Diabetes Dev Ctries 1-6. |
| [49] |
Ceriello A, De Nigris V, Prattichizzo F (2020) Why is hyperglycaemia worsening COVID-19 and its prognosis? Diabetes Obes Metab 22: 1951-1952. doi: 10.1111/dom.14098
|
| [50] |
Lee MH, Wong C, Ng CH, et al. (2021) Effects of hyperglycaemia on complications of COVID-19: a meta-analysis of observational studies. Diabetes Obes Metab 23: 287-289. doi: 10.1111/dom.14184
|
| [51] |
Singh AK, Singh R (2020) Hyperglycemia without diabetes and new-onset diabetes are both associated with poorer outcomes in COVID-19. Diabetes Res Clin Pract 167: 108382. doi: 10.1016/j.diabres.2020.108382
|
| [52] |
Carrasco-Sánchez FJ, López-Carmona MD, Martínez-Marcos FJ, et al. (2021) Admission hyperglycaemia as a predictor of mortality in patients hospitalized with COVID-19 regardless of diabetes status: data from the Spanish SEMI-COVID-19 Registry. Ann Med 53: 103-116. doi: 10.1080/07853890.2020.1836566
|
| [53] |
Wang S, Ma P, Zhang S, et al. (2020) Fasting blood glucose at admission is an independent predictor for 28-day mortality in patients with COVID-19 without previous diagnosis of diabetes: a multi-centre retrospective study. Diabetologia 63: 2102-2111. doi: 10.1007/s00125-020-05209-1
|
| [54] |
Li H, Tian S, Chen T, et al. (2020) Newly diagnosed diabetes is associated with a higher risk of mortality than known diabetes in hospitalized patients with COVID-19. Diabetes Obes Metab 22: 1897-1906. doi: 10.1111/dom.14099
|
| [55] |
Lim S, Bae JH, Kwon HS, et al. (2021) COVID-19 and diabetes mellitus: from pathophysiology to clinical management. Nat Rev Endocrinol 17: 11-30. doi: 10.1038/s41574-020-00435-4
|
| [56] |
Abu-Farha M, Al-Mulla F, Thanaraj TA, et al. (2020) Impact of diabetes in patients diagnosed with COVID-19. Front Immunol 11: 576818. doi: 10.3389/fimmu.2020.576818
|
| [57] | Yaribeygi H, Sathyapalan T, Jamialahmadi T, et al. (2020) The impact of diabetes mellitus in COVID-19: a mechanistic review of molecular interactions. J Diabetes Res 2020: 5436832. |
| [58] |
Singh AK, Gupta R, Ghosh A, et al. (2020) Diabetes in COVID-19: prevalence, pathophysiology, prognosis and practical considerations. Diabetes Metab Syndr 14: 303-310. doi: 10.1016/j.dsx.2020.04.004
|
| [59] |
Erener S (2020) Diabetes, infection risk and COVID-19. Mol Metab 39: 101044. doi: 10.1016/j.molmet.2020.101044
|
| [60] |
Azar WS, Njeim R, Fares AH, et al. (2020) COVID-19 and diabetes mellitus: how one pandemic worsens the other. Rev Endocr Metab Disord 21: 451-463. doi: 10.1007/s11154-020-09573-6
|
| [61] |
Banik GR, Alqahtani AS, Booy R, et al. (2016) Risk factors for severity and mortality in patients with MERS-CoV: analysis of publicly available data from Saudi Arabia. Virol Sin 31: 81-84. doi: 10.1007/s12250-015-3679-z
|
| [62] |
Kulcsar KA, Coleman CM, Beck SE, et al. (2019) Comorbid diabetes results in immune dysregulation and enhanced disease severity following MERS-CoV infection. JCI Insight 4: e131774. doi: 10.1172/jci.insight.131774
|
| [63] | Halaji M, Farahani A, Ranjbar R, et al. (2020) Emerging coronaviruses: first SARS, second MERS and third SARS-COV-2: epidemiological updates of COVID-19. Infez Med 28: 6-17. |
| [64] |
Muniyappa R, Gubbi S (2020) COVID-19 pandemic, coronaviruses, and diabetes mellitus. Am J Physiol Endocrinol Metab 318: E736-E741. doi: 10.1152/ajpendo.00124.2020
|
| [65] | Petersmann A, Nauck M, Müller-Wieland D, et al. (2018) Definition, classification and diagnosis of diabetes mellitus. Exp Clin Endocrinol Diabetes 127: S1-S7. |
| [66] |
Punthakee Z, Goldenberg R, et al. (2018) Definition, classification and diagnosis of diabetes, prediabetes and metabolic syndrome. Can J Diabetes 42: S10-S15. doi: 10.1016/j.jcjd.2017.10.003
|
| [67] |
Daryabor G, Atashzar MR, Kabelitz D, et al. (2020) The effects of type 2 diabetes mellitus on organ metabolism and the immune system. Front Immunol 11: 1582. doi: 10.3389/fimmu.2020.01582
|
| [68] |
Papatheodorou K, Banach M, Bekiari E, et al. (2018) Complications of diabetes 2017. J Diabetes Res 2018: 3086167. doi: 10.1155/2018/3086167
|
| [69] | Huang D, Refaat M, Mohammedi K, et al. (2017) Macrovascular complications in patients with diabetes and prediabetes. Biomed Res Int 2017: 7839101. |
| [70] |
Khalil H (2017) Diabetes microvascular complications – A clinical update. Diabetes Metab Syndr 11: S133-S139. doi: 10.1016/j.dsx.2016.12.022
|
| [71] |
Struijs JN, Baan CA, Schellevis FG, et al. (2006) Comorbidity in patients with diabetes mellitus: impact on medical health care utilization. BMC Health Serv Res 6: 84. doi: 10.1186/1472-6963-6-84
|
| [72] | Prattichizzo F, de Candia P, Nicolucci A, et al. (2021) Elevated HbA1c levels in pre-Covid-19 infection increases the risk of mortality: a sistematic review and meta-analysis. Diabetes Metab Res Rev e3476. |
| [73] |
Mirani M, Favacchio G, Carrone F, et al. (2020) Impact of comorbidities and glycemia at admission and dipeptidyl peptidase 4 inhibitors in patients with type 2 diabetes with covid-19: a case series from an academic hospital in lombardy, Italy. Diabetes Care 43: 3042-3049. doi: 10.2337/dc20-1340
|
| [74] |
Orioli L, Servais T, Belkhir L, et al. (2021) Clinical characteristics and short-term prognosis of in-patients with diabetes and COVID-19: a retrospective study from an academic center in Belgium. Diabetes Metab Syndr 15: 149-157. doi: 10.1016/j.dsx.2020.12.020
|
| [75] |
Sun Y, Guan X, Jia L, et al. (2021) Independent and combined effects of hypertension and diabetes on clinical outcomes in patients with COVID-19: a retrospective cohort study of Huoshen Mountain Hospital and Guanggu Fangcang Shelter Hospital. J Clin Hypertens 23: 218-231. doi: 10.1111/jch.14146
|
| [76] |
Vieira E, Mirizio GG, Barin GR, et al. (2020) Clock genes, inflammation and the immune system—Implications for diabetes, obesity and neurodegenerative diseases. Int J Mol Sci 21: 9743. doi: 10.3390/ijms21249743
|
| [77] | Zhou T, Hu Z, Yang S, et al. (2018) Role of adaptive and innate immunity in type 2 diabetes mellitus. J Diabetes Res 2018: 7457269. |
| [78] |
Hodgson K, Morris J, Bridson T, et al. (2015) Immunological mechanisms contributing to the double burden of diabetes and intracellular bacterial infections. Immunology 144: 171-185. doi: 10.1111/imm.12394
|
| [79] |
Alzaid F, Julla J, Diedisheim M, et al. (2020) Monocytopenia, monocyte morphological anomalies and hyperinflammation characterise severe COVID-19 in type 2 diabetes. EMBO Mol Med 12: e13038. doi: 10.15252/emmm.202013038
|
| [80] |
De Candia P, Prattichizzo F, Garavelli S, et al. (2019) Type 2 diabetes: how much of an autoimmune disease? Front Endocrinol (Lausanne) 10: 451. doi: 10.3389/fendo.2019.00451
|
| [81] |
Prattichizzo F, Giuliani A, Sabbatinelli J, et al. (2020) Prevalence of residual inflammatory risk and associated clinical variables in patients with type 2 diabetes. Diabetes Obes Metab 22: 1696-1700. doi: 10.1111/dom.14081
|
| [82] |
Maedler K, Spinas GA, Lehmann R, et al. (2001) Glucose induces β-Cell apoptosis via upregulation of the Fas receptor in human islets. Diabetes 50: 1683-1690. doi: 10.2337/diabetes.50.8.1683
|
| [83] |
Maedler K, Sergeev P, Ris F, et al. (2002) Glucose-induced β cell production of IL-1β contributes to glucotoxicity in human pancreatic islets. J Clin Invest 110: 851-860. doi: 10.1172/JCI200215318
|
| [84] | Zhou W, Ye S, Wang W, et al. (2020) Clinical features of COVID-19 patients with diabetes and secondary hyperglycemia. J Diabetes Res 2020: 3918723. |
| [85] |
Roganović J (2021) Downregulation of microRNA-146a in diabetes, obesity and hypertension may contribute to severe COVID-19. Med Hypotheses 146: 110448. doi: 10.1016/j.mehy.2020.110448
|
| [86] |
Gou L, Xiang M, Ran X, et al. (2021) Hyperosmolarity deserves more attention in critically Ill COVID-19 patients with diabetes: a cohort-based study. Diabetes Metab Syndr Obes Targets Ther 14: 47-58. doi: 10.2147/DMSO.S284148
|
| [87] |
Zheng M, Wang X, Guo H, et al. (2021) The cytokine profiles and immune response are increased in COVID-19 patients with type 2 diabetes mellitus. J Diabetes Res 2021: 9526701. doi: 10.1155/2021/9526701
|
| [88] | Guo W, Li M, Dong Y, et al. (2020) Diabetes is a risk factor for the progression and prognosis of COVID-19. Diabetes Metab Res Rev e3319. |
| [89] |
Cheng Y, Yue L, Wang Z, et al. (2021) Hyperglycemia associated with lymphopenia and disease severity of COVID-19 in type 2 diabetes mellitus. J Diabetes Complications 35: 107809. doi: 10.1016/j.jdiacomp.2020.107809
|
| [90] | Sun Y, Zhao R, Hu Z, et al. (2020) Differences in the clinical and hematological characteristics of COVID-19 patients with and without type 2 diabetes. J Diabetes Res 2020: 1038585. |
| [91] |
Zhang Y, Li H, Zhang J, et al. (2020) The clinical characteristics and outcomes of patients with diabetes and secondary hyperglycaemia with coronavirus disease 2019: a single-centre, retrospective, observational study in Wuhan. Diabetes Obes Metab 22: 1443-1454. doi: 10.1111/dom.14086
|
| [92] |
Ulhaq ZS, Soraya GV (2020) Interleukin-6 as a potential biomarker of COVID-19 progression. Médecine Mal Infect 50: 382-383. doi: 10.1016/j.medmal.2020.04.002
|
| [93] |
Scheller J, Chalaris A, Schmidt-Arras D, et al. (2011) The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim Biophys Acta 1813: 878-888. doi: 10.1016/j.bbamcr.2011.01.034
|
| [94] | Muangchan C, Pope JE (2013) The significance of interleukin-6 and C-reactive protein in systemic sclerosis: a systematic literature review. Clin Exp Rheumatol 31: 122-134. |
| [95] |
Mishra Y, Pathak BK, Mohakuda SS, et al. (2020) Relation of D-dimer levels of COVID-19 patients with diabetes mellitus. Diabetes Metab Syndr 14: 1927-1930. doi: 10.1016/j.dsx.2020.09.035
|
| [96] |
Rostami M, Mansouritorghabeh H (2020) D-dimer level in COVID-19 infection: a systematic review. Expert Rev Hematol 13: 1265-1275. doi: 10.1080/17474086.2020.1831383
|
| [97] |
Mei H, Luo L, Hu Y (2020) Thrombocytopenia and thrombosis in hospitalized patients with COVID-19. J Hematol Oncol 13: 161. doi: 10.1186/s13045-020-01003-z
|
| [98] |
Zahran AM, El-Badawy O, Mohamad IL, et al. (2018) Platelet activation and platelet–leukocyte aggregates in type I diabetes mellitus. Clin Appl Thromb Hemost 24: 230S-239S. doi: 10.1177/1076029618805861
|
| [99] |
Kaur R, Kaur M, Singh J (2018) Endothelial dysfunction and platelet hyperactivity in type 2 diabetes mellitus: molecular insights and therapeutic strategies. Cardiovasc Diabetol 17: 121. doi: 10.1186/s12933-018-0763-3
|
| [100] |
Shang J, Wang Q, Zhang H, et al. (2021) The relationship between diabetes mellitus and COVID-19 prognosis: a retrospective cohort study in Wuhan, China. Am J Med 134: e6-e14. doi: 10.1016/j.amjmed.2020.05.033
|
| [101] | Guo T, Shen Q, Ouyang X, et al. (2021) Clinical findings in diabetes mellitus patients with COVID-19. J Diabetes Res 2021: 7830136. |
| [102] |
Al-Salameh A, Lanoix JP, Bennis Y, et al. (2021) Characteristics and outcomes of COVID-19 in hospitalized patients with and without diabetes. Diabetes Metab Res Rev 37: e3388. doi: 10.1002/dmrr.3388
|
| [103] | Chen X, Hu W, Ling J, et al. (2020) Hypertension and diabetes delay the viral clearance in COVID-19 patients. medRxiv . |
| [104] |
Clarke NE, Turner AJ (2012) Angiotensin-converting enzyme 2: the first decade. Int J Hypertens 2012: 307315. doi: 10.1155/2012/307315
|
| [105] |
Warner FJ, Smith AI, Hooper NM, et al. (2004) Angiotensin-converting enzyme-2: a molecular and cellular perspective. Cell Mol Life Sci 61: 2704-2713. doi: 10.1007/s00018-004-4240-7
|
| [106] |
Santos RAS, Ferreira AJ, Simões E, Silva AC (2008) Recent advances in the angiotensin-converting enzyme 2-angiotensin(1-7)-Mas axis. Exp Physiol 93: 519-527. doi: 10.1113/expphysiol.2008.042002
|
| [107] |
Santos RAS, Ferreira AJ, Verano-Braga T, et al. (2013) Angiotensin-converting enzyme 2, angiotensin-(1-7) and Mas: new players of the renin-angiotensin system. J Endocrinol 216: R1-R17. doi: 10.1530/JOE-12-0341
|
| [108] |
Hashimoto T, Perlot T, Rehman A, et al. (2012) ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 487: 477-481. doi: 10.1038/nature11228
|
| [109] |
Perlot T, Penninger JM (2013) ACE2 – From the renin-angiotensin system to gut microbiota and malnutrition. Microbes Infect 15: 866-873. doi: 10.1016/j.micinf.2013.08.003
|
| [110] |
Hamming I, Timens W, Bulthuis MLC, et al. (2004) Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J Pathol 203: 631-637. doi: 10.1002/path.1570
|
| [111] |
Tikellis C, Johnston CI, Forbes JM, et al. (2003) Characterization of renal angiotensin-converting enzyme 2 in diabetic nephropathy. Hypertension 41: 392-397. doi: 10.1161/01.HYP.0000060689.38912.CB
|
| [112] |
Xia H, Lazartigues E (2008) Angiotensin-converting enzyme 2 in the brain: properties and future directions. J Neurochem 107: 1482-1494. doi: 10.1111/j.1471-4159.2008.05723.x
|
| [113] |
Xu H, Zhong L, Deng J, et al. (2020) High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int J Oral Sci 12: 8. doi: 10.1038/s41368-020-0074-x
|
| [114] |
To KF, Lo AWI (2004) Exploring the pathogenesis of severe acute respiratory syndrome (SARS): The tissue distribution of the coronavirus (SARS-CoV) and its putative receptor, angiotensin-converting enzyme 2 (ACE2). J Pathol 203: 740-743. doi: 10.1002/path.1597
|
| [115] |
Batlle D, Wysocki J, Satchell K (2020) Soluble angiotensin-converting enzyme 2: a potential approach for coronavirus infection therapy? Clin Sci 134: 543-545. doi: 10.1042/CS20200163
|
| [116] |
Ni W, Yang X, Yang D, et al. (2020) Role of angiotensin-converting enzyme 2 (ACE2) in COVID-19. Crit Care 24: 422. doi: 10.1186/s13054-020-03120-0
|
| [117] |
Zhang H, Penninger JM, Li Y, et al. (2020) Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: molecular mechanisms and potential therapeutic target. Intensive Care Med 46: 586-590. doi: 10.1007/s00134-020-05985-9
|
| [118] |
Shang J, Wan Y, Luo C, et al. (2020) Cell entry mechanisms of SARS-CoV-2. Proc Natl Acad Sci U S A 117: 11727-11734. doi: 10.1073/pnas.2003138117
|
| [119] |
Valencia I, Peiró C, Lorenzo Ó, et al. (2020) DPP4 and ACE2 in diabetes and COVID-19: therapeutic targets for cardiovascular complications? Front Pharmacol 11: 1161. doi: 10.3389/fphar.2020.01161
|
| [120] |
Bourgonje AR, Abdulle AE, Timens W, et al. (2020) Angiotensin-converting enzyme 2 (ACE2), SARS-CoV-2 and the pathophysiology of coronavirus disease 2019 (COVID-19). J Pathol 251: 228-248. doi: 10.1002/path.5471
|
| [121] | Chen J, Jiang Q, Xia X, et al. (2020) Individual variation of the SARS-CoV-2 receptor ACE2 gene expression and regulation. Aging Cell 19: e13168. |
| [122] |
Mizuiri S, Hemmi H, Arita M, et al. (2008) Expression of ACE and ACE2 in individuals with diabetic kidney disease and healthy controls. Am J Kidney Dis 51: 613-623. doi: 10.1053/j.ajkd.2007.11.022
|
| [123] |
Reich HN, Oudit GY, Penninger JM, et al. (2008) Decreased glomerular and tubular expression of ACE2 in patients with type 2 diabetes and kidney disease. Kidney Int 74: 1610-1616. doi: 10.1038/ki.2008.497
|
| [124] |
Gilbert RE, Caldwell L, Misra PS, et al. (2021) Overexpression of the severe acute respiratory syndrome coronavirus-2 receptor, angiotensin-converting enzyme 2, in diabetic kidney disease: implications for kidney injury in novel coronavirus disease 2019. Can J Diabetes 45: 162-166.e1. doi: 10.1016/j.jcjd.2020.07.003
|
| [125] |
Soldo J, Heni M, Königsrainer A, et al. (2020) Increased hepatic ace2 expression in nafl and diabetes-a risk for covid-19 patients? Diabetes Care 43: e134-e136. doi: 10.2337/dc20-1458
|
| [126] |
Wijnant SRA, Jacobs M, Van Eeckhoutte HP, et al. (2020) Expression of ACE2, the SARS-CoV-2 receptor, in lung tissue of patients with type 2 diabetes. Diabetes 69: 2691-2699. doi: 10.2337/db20-0669
|
| [127] |
Parit R, Jayavel S (2021) Association of ACE inhibitors and angiotensin type II blockers with ACE2 overexpression in COVID-19 comorbidities: a pathway-based analytical study. Eur J Pharmacol 896: 173899. doi: 10.1016/j.ejphar.2021.173899
|
| [128] |
Pal R, Bhadada SK (2020) COVID-19 and diabetes mellitus: an unholy interaction of two pandemics. Diabetes Metab Syndr 14: 513-517. doi: 10.1016/j.dsx.2020.04.049
|
| [129] |
Menon R, Otto EA, Sealfon R, et al. (2020) SARS-CoV-2 receptor networks in diabetic and COVID-19–associated kidney disease. Kidney Int 98: 1502-1518. doi: 10.1016/j.kint.2020.09.015
|
| [130] |
Batchu SN, Kaur H, Yerra VG, et al. (2021) Lung and kidney ACE2 and TMPRSS2 in renin-angiotensin system blocker-treated comorbid diabetic mice mimicking host factors that have been linked to severe COVID-19. Diabetes 70: 759-771. doi: 10.2337/db20-0765
|
| [131] |
Sriram K, Insel PA (2020) Risks of ACE inhibitor and ARB usage in COVID-19: evaluating the evidence. Clin Pharmacol Ther 108: 236-241. doi: 10.1002/cpt.1863
|
| [132] |
Burrell LM, Harrap SB, Velkoska E, et al. (2013) The ACE2 gene: its potential as a functional candidate for cardiovascular disease. Clin Sci 124: 65-76. doi: 10.1042/CS20120269
|
| [133] |
Meng N, Zhang Y, Ma J, et al. (2015) Association of polymorphisms of angiotensin i converting enzyme 2 with retinopathy in type 2 diabetes mellitus among Chinese individuals. Eye 29: 266-271. doi: 10.1038/eye.2014.254
|
| [134] |
Fröjdö S, Sjölind L, Parkkonen M, et al. (2005) Polymorphisms in the gene encoding angiotensin I converting enzyme 2 and diabetic nephropathy. Diabetologia 48: 2278-2281. doi: 10.1007/s00125-005-1955-4
|
| [135] |
Currie D, McKnight AJ, Patterson CC, et al. (2010) Investigation of ACE, ACE2 and AGTR1 genes for association with nephropathy in type 1 diabetes mellitus. Diabet Med 27: 1188-1194. doi: 10.1111/j.1464-5491.2010.03097.x
|
| [136] | Chaoxin J, Daili S, Yanxin H, et al. (2013) The influence of angiotensin-converting enzyme 2 gene polymorphisms on type 2 diabetes mellitus and coronary heart disease. Eur Rev Med Pharmacol Sci 17: 2654-2659. |
| [137] |
Patel SK, Wai B, Ord M, et al. (2012) Association of ACE2 genetic variants with blood pressure, left ventricular mass, and cardiac function in caucasians with type 2 diabetes. Am J Hypertens 25: 216-222. doi: 10.1038/ajh.2011.188
|
| [138] |
Sayed S (2021) COVID-19 and diabetes; Possible role of polymorphism and rise of telemedicine. Prim Care Diabetes 15: 4-9. doi: 10.1016/j.pcd.2020.08.018
|
| [139] |
Calcagnile M, Forgez P, Iannelli A, et al. (2021) Molecular docking simulation reveals ACE2 polymorphisms that may increase the affinity of ACE2 with the SARS-CoV-2 spike protein. Biochimie 180: 143-148. doi: 10.1016/j.biochi.2020.11.004
|
| [140] | Calcagnile M, Forgez P, Iannelli A, et al. (2020) ACE2 polymorphisms and individual susceptibility to SARS-CoV-2 infection: insights from an in silico study. bioRxiv . |
| [141] |
Hamet P, Pausova Z, Attaoua R, et al. (2021) SARS-COV-2 receptor ACE2 gene is associated with hypertension and severity of COVID 19: interaction with sex, obesity and smoking. Am J Hypertens 34: 367-376. doi: 10.1093/ajh/hpaa223
|
| [142] |
Choudhary S, Sreenivasulu K, Mitra P, et al. (2020) Role of genetic variants and gene expression in the susceptibility and severity of COVID-19. Ann Lab Med 41: 129-138. doi: 10.3343/alm.2021.41.2.129
|
| [143] |
Paniri A, Hosseini MM, Moballegh-Eslam M, et al. (2021) Comprehensive in silico identification of impacts of ACE2 SNPs on COVID-19 susceptibility in different populations. Gene Rep 22: 100979. doi: 10.1016/j.genrep.2020.100979
|
| [144] |
Wee AKH (2021) COVID-19's toll on the elderly and those with diabetes mellitus – Is vitamin B12 deficiency an accomplice? Med Hypotheses 146: 110374. doi: 10.1016/j.mehy.2020.110374
|
| [145] |
Gonçalves SEAB, Gonçalves TJM, Guarnieri A, et al. (2021) Association between thiamine deficiency and hyperlactatemia among critically ill patients with diabetes infected by SARS-CoV-2. J Diabetes 13: 413-419. doi: 10.1111/1753-0407.13156
|
| [146] |
Rhodes JM, Subramanian S, Laird E, et al. (2021) Perspective: vitamin D deficiency and COVID-19 severity – plausibly linked by latitude, ethnicity, impacts on cytokines, ACE2 and thrombosis. J Intern Med 289: 97-115. doi: 10.1111/joim.13149
|
| [147] |
Tort AR, Montelongo EAM, Cuazitl AM, et al. (2020) La deficiencia de vitamina D es un factor de riesgo de mortalidad en pacientes con COVID-19. Rev Sanid Milit 74: 106-113. doi: 10.35366/93773
|
| [148] |
Biesalski HK (2020) Vitamin D deficiency and co-morbidities in COVID-19 patients – A fatal relationship? NFS J 20: 10. doi: 10.1016/j.nfs.2020.06.001
|
| [149] |
Berridge MJ (2017) Vitamin D deficiency and diabetes. Biochem J 474: 1321-1332. doi: 10.1042/BCJ20170042
|
| [150] |
Luo BA, Gao F, Qin LL (2017) The association between vitamin D deficiency and diabetic retinopathy in type 2 diabetes: A meta-analysis of observational studies. Nutrients 9: 307. doi: 10.3390/nu9030307
|
| [151] |
Singh SK, Jain R, Singh S (2020) Vitamin D deficiency in patients with diabetes and COVID-19 infection. Diabetes Metab Syndr 14: 1033-1035. doi: 10.1016/j.dsx.2020.06.071
|
| [152] |
Jin S, Hu W (2021) Severity of COVID-19 and treatment strategy for patient with diabetes. Front Endocrinol (Lausanne) 12: 602735. doi: 10.3389/fendo.2021.602735
|
| [153] |
Ceriello A, Prattichizzo F (2021) Pharmacological management of COVID-19 in type 2 diabetes. J Diabetes Complications 35: 107927. doi: 10.1016/j.jdiacomp.2021.107927
|
| [154] |
Deng F, Gao D, Ma X, et al. (2021) Corticosteroids in diabetes patients infected with COVID-19. Ir J Med Sci 190: 29-31. doi: 10.1007/s11845-020-02287-3
|
| [155] |
Stoian AP, Catrinoiu D, Rizzo M, et al. (2021) Hydroxychloroquine, COVID-19 and diabetes. Why it is a different story. Diabetes Metab Res Rev 37: e3379. doi: 10.1002/dmrr.3379
|
| [156] |
Khunti K, Knighton P, Zaccardi F, et al. (2021) Prescription of glucose-lowering therapies and risk of COVID-19 mortality in people with type 2 diabetes : a nationwide observational study in England. Lancet Diabetes Endocrinol 9: 293-303. doi: 10.1016/S2213-8587(21)00050-4
|
| [157] |
Gregory JM, Slaughter JC, Duffus SH, et al. (2021) COVID-19 severity is tripled in the diabetes community: a prospective analysis of the pandemic's impact in type 1 and type 2 diabetes. Diabetes Care 44: 526-532. doi: 10.2337/dc20-2260
|
| [158] |
Fisher L, Polonsky W, Asuni A, et al. (2020) The early impact of the COVID-19 pandemic on adults with type 1 or type 2 diabetes: a national cohort study. J Diabetes Complications 34: 107748. doi: 10.1016/j.jdiacomp.2020.107748
|
| [159] |
Ruissen MM, Regeer H, Landstra CP, et al. (2021) Increased stress, weight gain and less exercise in relation to glycemic control in people with type 1 and type 2 diabetes during the COVID-19 pandemic. BMJ Open Diabetes Res Care 9: e002035. doi: 10.1136/bmjdrc-2020-002035
|
| [160] |
Holman N, Knighton P, Kar P, et al. (2020) Risk factors for COVID-19-related mortality in people with type 1 and type 2 diabetes in England: a population-based cohort study. Lancet Diabetes Endocrinol 8: 823-833. doi: 10.1016/S2213-8587(20)30271-0
|
| [161] |
Beliard K, Ebekozien O, Demeterco-Berggren C, et al. (2021) Increased DKA at presentation among newly diagnosed type 1 diabetes patients with or without COVID-19: data from a multi-site surveillance registry. J Diabetes 13: 270-272. doi: 10.1111/1753-0407.13141
|
| [162] |
Vamvini M, Lioutas VA, Middelbeek RJW (2020) Characteristics and diabetes control in adults with type 1 diabetes admitted with covid-19 infection. Diabetes Care 43: e120-e122. doi: 10.2337/dc20-1540
|
| [163] |
Al Hayek AA, Robert AA, Alotaibi ZK, et al. (2020) Clinical characteristics of hospitalized and home isolated COVID-19 patients with type 1 diabetes. Diabetes Metab Syndr 14: 1841-1845. doi: 10.1016/j.dsx.2020.09.013
|
| [164] |
Unsworth R, Wallace S, Oliver NS, et al. (2020) New-onset type 1 diabetes in children during COVID-19: multicenter regional findings in the U.K.. Diabetes Care 43: e170-e171. doi: 10.2337/dc20-1551
|
| [165] |
Suwanwongse K, Shabarek N (2021) Newly diagnosed diabetes mellitus, DKA, and COVID-19: causality or coincidence? A report of three cases. J Med Virol 93: 1150-1153. doi: 10.1002/jmv.26339
|
| [166] |
Dimeglio LA, Albanese-O'neill A, Muñoz CE, et al. (2020) COVID-19 and children with diabetesdupdates, unknowns, and next steps: first, do no extrapolation. Diabetes Care 43: 2631-2634. doi: 10.2337/dci20-0044
|
| [167] |
Cameron FJ, Scratch SE, Nadebaum C, et al. (2014) Neurological consequences of diabetic ketoacidosis at initial presentation of type 1 diabetes in a prospective cohort study of children. Diabetes Care 37: 1554-1562. doi: 10.2337/dc13-1904
|
| [168] |
Kamrath C, Mönkemöller K, Biester T, et al. (2020) Ketoacidosis in children and adolescents with newly diagnosed type 1 diabetes during the COVID-19 pandemic in Germany. JAMA 324: 801-804. doi: 10.1001/jama.2020.13445
|
| [169] |
Rabbone I, Schiaffini R, Cherubini V, et al. (2020) Has COVID-19 delayed the diagnosis and worsened the presentation of type 1 diabetes in children? Diabetes Care 43: 2870-2872. doi: 10.2337/dc20-1321
|
| [170] |
Powers AC, Aronoff DM, Eckel RH (2021) COVID-19 vaccine prioritisation for type 1 and type 2 diabetes. Lancet Diabetes Endocrinol 9: 140-141. doi: 10.1016/S2213-8587(21)00017-6
|
| [171] |
Boddu SK, Aurangabadkar G, Kuchay MS (2020) New onset diabetes, type 1 diabetes and COVID-19. Diabetes Metab Syndr 14: 2211-2217. doi: 10.1016/j.dsx.2020.11.012
|
| [172] |
Kuchay MS, Reddy PK, Gagneja S, et al. (2020) Short term follow-up of patients presenting with acute onset diabetes and diabetic ketoacidosis during an episode of COVID-19. Diabetes Metab Syndr 14: 2039-2041. doi: 10.1016/j.dsx.2020.10.015
|
| [173] |
Marchand L, Pecquet M, Luyton C (2020) Type 1 diabetes onset triggered by COVID-19. Acta Diabetol 57: 1265-1266. doi: 10.1007/s00592-020-01570-0
|
| [174] |
Caruso P, Longo M, Esposito K, et al. (2020) Type 1 diabetes triggered by covid-19 pandemic: a potential outbreak? Diabetes Res Clin Pract 164: 108219. doi: 10.1016/j.diabres.2020.108219
|
| [175] |
Goyal H, Sachdeva S, Perisetti A, et al. (2021) Hyperlipasemia and potential pancreatic injury patterns in COVID-19: a marker of severity or innocent bystander? Gastroenterology 160: 946-948.e2. doi: 10.1053/j.gastro.2020.10.037
|
| [176] |
Patnaik RNK, Gogia A, Kakar A (2020) Acute pancreatic injury induced by COVID-19. IDCases 22: e00959. doi: 10.1016/j.idcr.2020.e00959
|
| [177] |
Bruno G, Fabrizio C, Santoro CR, et al. (2021) Pancreatic injury in the course of coronavirus disease 2019: a not-so-rare occurrence. J Med Virol 93: 74-75. doi: 10.1002/jmv.26134
|
| [178] |
Ghosh A, Gupta V, Misra A (2020) COVID19 induced acute pancreatitis and pancreatic necrosis in a patient with type 2 diabetes. Diabetes Metab Syndr 14: 2097-2098. doi: 10.1016/j.dsx.2020.10.008
|
| [179] |
Alharmi RAR, Fateel T, Sayed Adnan J, et al. (2021) Acute pancreatitis in a patient with COVID-19. BMJ Case Rep 14: e239656. doi: 10.1136/bcr-2020-239656
|
| [180] |
Wang F, Wang H, Fan J, et al. (2020) Pancreatic injury patterns in patients with coronavirus disease 19 pneumonia. Gastroenterology 159: 367-370. doi: 10.1053/j.gastro.2020.03.055
|
| [181] |
Pribadi RR, Simadibrata M (2021) Increased serum amylase and/or lipase in coronavirus disease 2019 (COVID-19) patients: is it really pancreatic injury? JGH Open 5: 190-192. doi: 10.1002/jgh3.12436
|
| [182] |
Coate KC, Cha J, Shrestha S, et al. (2020) SARS-CoV-2 cell entry factors ACE2 and TMPRSS2 are expressed in the microvasculature and ducts of human pancreas but are not enriched in β cells. Cell Metab 32: 1028-1040.e4. doi: 10.1016/j.cmet.2020.11.006
|
| [183] |
Kusmartseva I, Wu W, Syed F, et al. (2020) Expression of SARS-CoV-2 entry factors in the pancreas of normal organ donors and individuals with COVID-19. Cell Metab 32: 1041-1051.e6. doi: 10.1016/j.cmet.2020.11.005
|
| [184] |
Müller JA, Groß R, Conzelmann C, et al. (2021) SARS-CoV-2 infects and replicates in cells of the human endocrine and exocrine pancreas. Nat Metab 3: 149-165. doi: 10.1038/s42255-021-00347-1
|
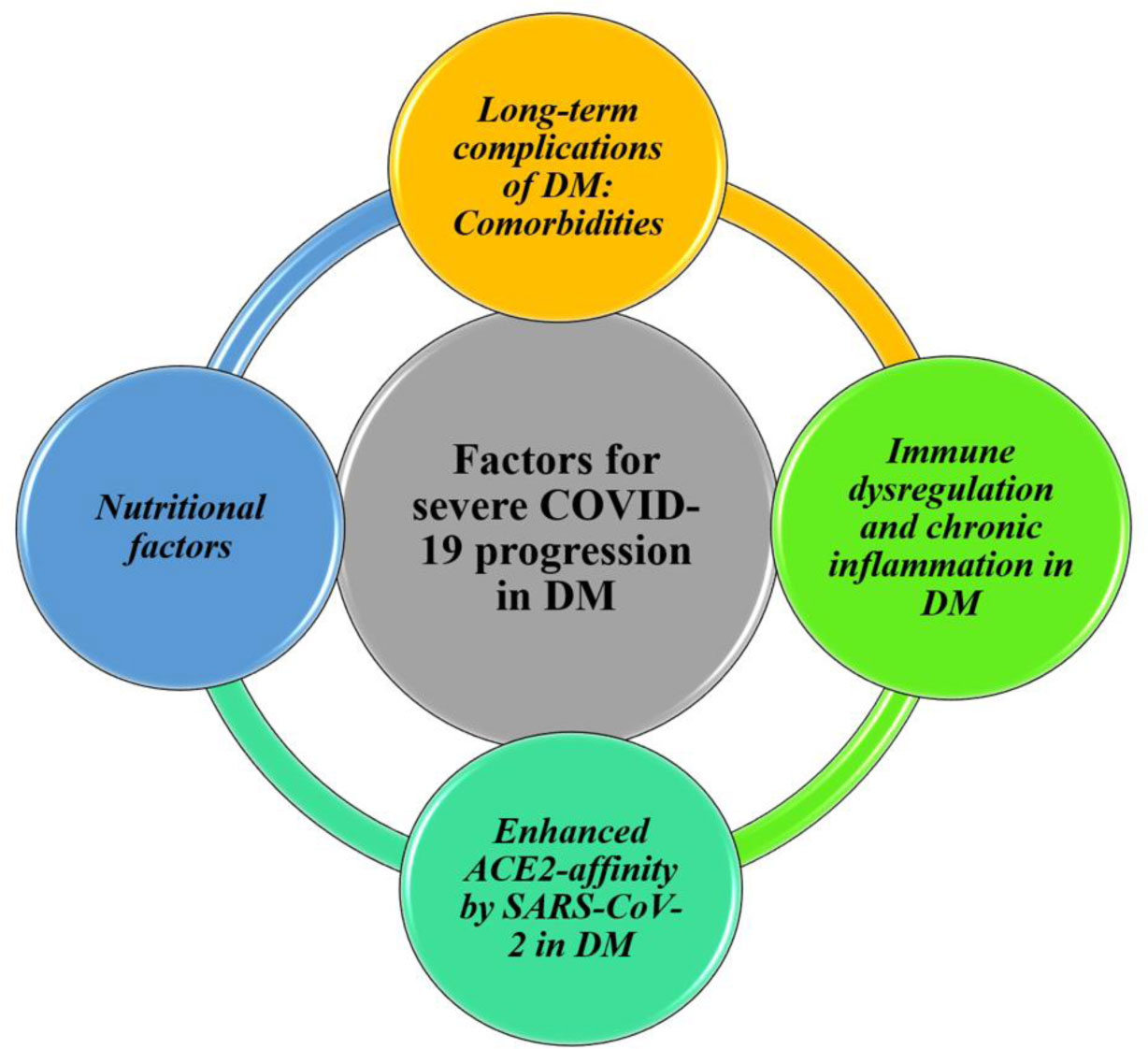
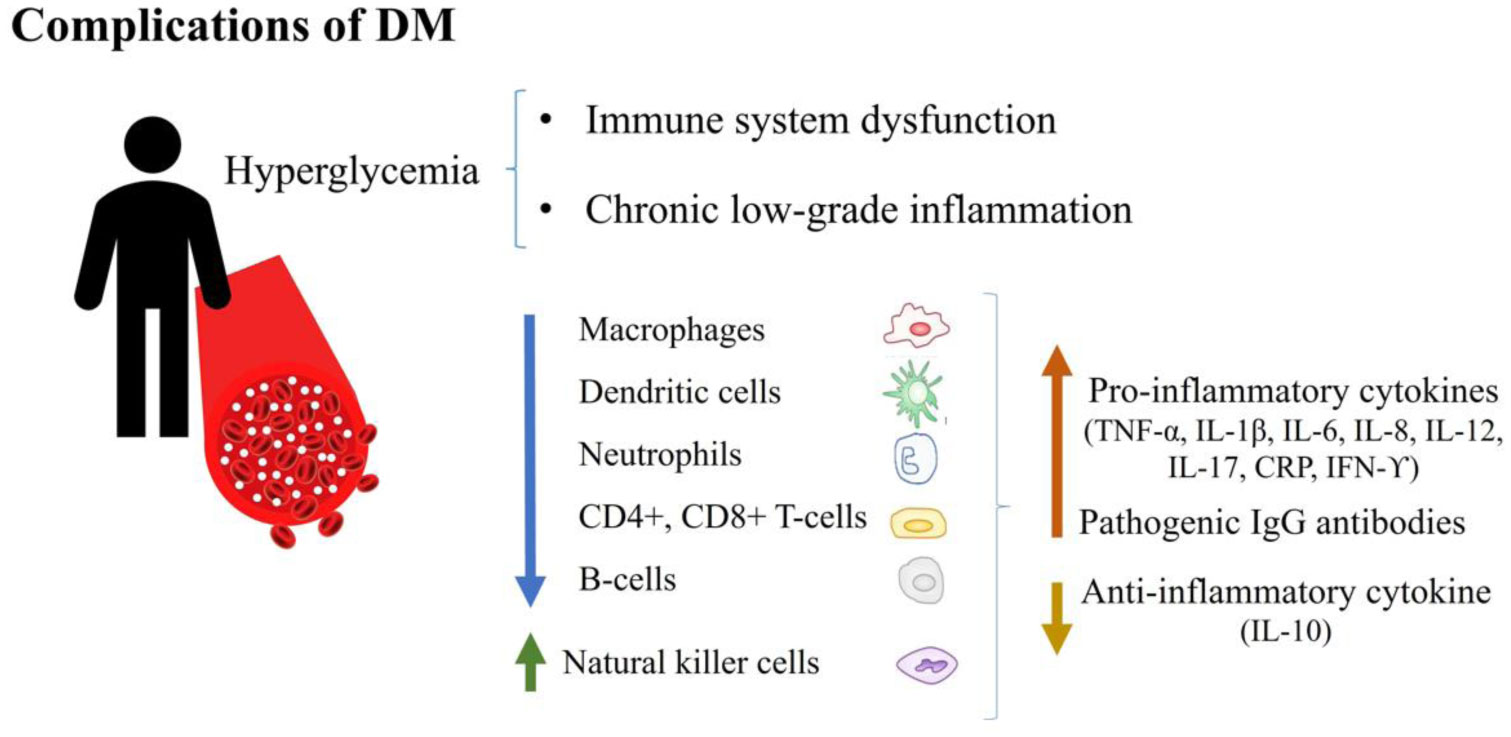
Figures(2)

María D Figueroa-Pizano, Alma C Campa-Mada, Elizabeth Carvajal-Millan, Karla G Martinez-Robinson, Agustin Rascon Chu. The underlying mechanisms for severe COVID-19 progression in people with diabetes mellitus: a critical review[J]. AIMS Public Health, 2021, 8(4): 720-742. doi: 10.3934/publichealth.2021057







 DownLoad:
DownLoad: