Citation: Wenpeng You, Maciej Henneberg. Cereal Crops Are not Created Equal: Wheat Consumption Associated with Obesity Prevalence Globally and Regionally[J]. AIMS Public Health, 2016, 3(2): 313-328. doi: 10.3934/publichealth.2016.2.313

| [1] | Rokholm B, Baker JL, Sørensen TIA. (1999) The levelling off of the obesity epidemic since the year — a review of evidence and perspectives. Oxford, UK2010. p. 835-46. |
| [2] | Popkin BM, Adair LS, Ng SW. (2012) Global nutrition transition and the pandemic of obesity in developing countries. Nutrition Reviews 70(1): 3-21. |
| [3] | Stevens G, Singh G, Lu Y, Danaei G, Lin J, Finucane MM, et al. (2012) National, regional, and global trends in adult overweight and obesity prevalences. Population Health Metrics 10. |
| [4] | WHO. Obesity: Preventing and Managing the Global Epidemic. Geneva World Health Organization 2000.2004. |
| [5] | Nguyen DM, El-Serag H. (2010) The Epidemiology of Obesity. Gastroenterol Clin North Am 39(1): 1-7. |
| [6] | You W, Henneberg M. (2016) Meat consumption providing a surplus energy in modern diet contributes to obesity prevalence: an ecological analysis. BMC Nutrition 2(1). |
| [7] | Roccisano D, Henneberg M. (2012) Soy Consumption and Obesity. Food and Nutrition Sciences 03(02): 260-6. |
| [8] |
Weeratunga P, Jayasinghe S, Perera Y, Jayasena G, Jayasinghe S. (2014) Per capita sugar consumption and prevalence of diabetes mellitus – global and regional associations. BMC Public Health 14: 186-91. doi: 10.1186/1471-2458-14-186

|
| [9] | Basu S, Yoffe P, Hills N, Lustig RH. (2013) The relationship of sugar to population-level diabetes prevalence: an econometric analysis of repeated cross-sectional data. PloS one 8(2):e57873: 1-8. |
| [10] | Kawada T. (2012) Physical activity, obesity and insulin resistance. International Journal of Cardiology 159(3): 237-8. |
| [11] | Jakicic JM, Davis KK. (2011) Obesity and Physical Activity. Psychiatric Clinics of North America 34(4): 829-40. |
| [12] | Henneberg M, Grantham J. (2014) Obesity - a natural consequence of human evolution. Anthropological Review 77(1): 1-10. |
| [13] | Moleres A, Martinez J, Marti A. (2013) Genetics of Obesity. Curr Obes Rep 2(1): 23-31. |
| [14] | Slavin JL, Martini MC, Jacobs DR, Marquart L. (1999) Plausible mechanisms for the protectiveness of whole grains. The American Journal of Clinical Nutrition 70(3 Suppl):459S. |
| [15] | Cancer Council Australia. Position statement - Fibre, wholegrain cereals and cancer - National Cancer Prevention Policy. Available from: http://wiki.cancer.org.au. |
| [16] | De La Hunty A, Ashwell M. (2007) Are people who regularly eat breakfast cereals slimmer than those who don’t? A systematic review of the evidence. Oxford, UK2007. p. 118-28. |
| [17] | de la Hunty A, Gibson S, Ashwell M. (2013) Does Regular Breakfast Cereal Consumption Help Children and Adolescents Stay Slimmer? A Systematic Review and Meta- Analysis. Obesity Facts 6(1): 70-85. |
| [18] | Bazzano LA, Song Y, Bubes V, Good CK, Manson JE, Liu S. (2005) Dietary intake of whole and refined grain breakfast cereals and weight gain in men. Obesity Research 13(11): 1952. |
| [19] | van de Vijver LPL, van den Bosch LMC, van den Brandt PA, Goldbohm RA. (2009) Whole- grain consumption, dietary fibre intake and body mass index in the Netherlands cohort study. European Journal of Clinical Nutrition 63(1): 31. |
| [20] | Cho SS, Qi L, Fahey GC, Jr., Klurfeld DM. (2013) Consumption of cereal fiber, mixtures of whole grains and bran, and whole grains and risk reduction in type 2 diabetes, obesity, and cardiovascular disease. The American Journal of Clinical Nutrition 98(2): 594-619. |
| [21] | Ye EQ, Chacko SA, Chou EL, Kugizaki M, Liu S. (2012) Greater whole-grain intake is associated with lower risk of type 2 diabetes, cardiovascular disease, and weight gain.(Nutritional Epidemiology)(Author abstract)(Report). The Journal of Nutrition 142(7): 1304. |
| [22] | Slavin J, Jacobs D, Marquart L. (1997) Whole- grain consumption and chronic disease: Protective mechanisms. Nutr Cancer p. 14-21. |
| [23] | Jacobs D, Marquart L, Slavin J, Kushi LH. (1998) Whole- grain intake and cancer: An expanded review and meta-analysis. Nutr Cancer p. 85-96. |
| [24] |
Aune D, Chan DS, Lau R, Vieira R, Greenwood DC, Kampman E, et al. (2011) Dietary fibre, whole grains, and risk of colorectal cancer: systematic review and dose-response meta-analysis of prospective studies. BMJ 343:d6617. doi: 10.1136/bmj.d6617
|
| [25] | FAO. FAO plant production and protection series. Rome: Rome : FAO; 2002. |
| [26] | Brouns FJPH, van Buul VJ, Shewry PR. (2013) Does wheat make us fat and sick? Journal of Cereal Science 58(2): 209-15. |
| [27] | Shi Z, Taylor AW, Hu G, Gill T, Wittert GA. (2012) Rice intake, weight change and risk of the metabolic syndrome development among Chinese adults: the Jiangsu Nutrition Study (JIN). Asia Pacific Journal of Clinical Nutrition 21(1): 35. |
| [28] |
Zhang JG, Wang ZH, Wang HJ, Du WW, Su C, Zhang J, et al. (2015) Dietary patterns and their associations with general obesity and abdominal obesity among young Chinese women. European Journal of Clinical Nutrition 69: 1009-14. doi: 10.1038/ejcn.2015.8
|
| [29] | Davis WR. (2011) Wheat Belly: Lose the Wheat, Lose the Weight, and Find Your Path Back to Health. Rodale Books. |
| [30] | Jönsson T, Olsson S, Ahrén B, Bøg-Hansen T, Dole A, Lindeberg S. (2005) Agrarian diet and diseases of affluence—Do evolutionary novel dietary lectins cause leptin resistance? BMC Endocrine Disorders 5(1): 10. |
| [31] | Cheng J, Brar PS, Lee AR, Green PHR. (2010) Body mass index in celiac disease: beneficial effect of a gluten-free diet. Journal of Clinical Gastroenterology 44(4): 267-71. |
| [32] | Soares FL, de Oliveira Matoso R, Teixeira LG, Menezes Z, Pereira SS, Alves AC, et al. (2013) Gluten-free diet reduces adiposity, inflammation and insulin resistance associated with the induction of PPAR-alpha and PPAR-gamma expression. The Journal of Nutritional Biochemistry 24(6): 1105-11. |
| [33] | Kabbani TA, Goldberg A, Kelly CP, Pallav K, Tariq S, Peer A, et al. (2012) Body mass index and the risk of obesity in coeliac disease treated with the gluten-free diet. Aliment Pharmacol Ther 35(6): 723-9. |
| [34] | Hyman M. (2012) The Blood Sugar Solution: The Bestselling Programme for Preventing Diabetes, Losing Weight and Feeling Great: Hodder & Stoughton General Division. |
| [35] | Zhanga J, Wanga H, Wanga Y, Xue H, Wang Z, Dua W, et al. (2015) Dietary patterns and their associations with childhood obesity in China. British Journal of Nutrition 1-7. |
| [36] |
Hauner H, Bechthold A, Boeing H, Brönstrup A, Buyken A, Leschik-Bonnet E, et al. (2012) Evidence-Based Guideline of the German Nutrition Society: Carbohydrate Intake and Prevention of Nutrition-Related Diseases. Annals of Nutrition and Metabolism 60: 1-58. doi: 10.1159/000338102
|
| [37] | FAO. Food Balance Sheets. A Handbook. Rome: Food and Agriculture Organization, 2001. |
| [38] | Siervo M, Montagnese C, Mathers JC, Soroka KR, Stephan BCM, Wells JCK. (2014) Sugar consumption and global prevalence of obesity and hypertension: an ecological analysis. Public Health Nutrition 17(3): 587-96. |
| [39] | Basu S, Stuckler D, McKee M, Galea G. (2013) Nutritional determinants of worldwide diabetes: an econometric study of food markets and diabetes prevalence in 173 countries. Public Health Nutrition 16(1): 1-8. |
| [40] | Maskova E, Paulickova I, Rysova J, Gabrovska D. (2011) Evidence for Wheat, Rye, and Barley Presence in Gluten Free Foods by PCR Method - Comparison with Elisa Method. Czech Journal of Food Sciences 29(1): 45-50. |
| [41] |
Davis B, Wansink B. (2015) Fifty years of fat: news coverage of trends that predate obesity prevalence. BMC Public Health 15: 1-6. doi: 10.1186/1471-2458-15-1
|
| [42] | den Engelsen C, Gorter KJ, Salome PL, Rutten GE. (2013) Development of metabolic syndrome components in adults with a healthy obese phenotype: a 3-year follow-up. Obesity 21(5): 1025-30. |
| [43] | Trøseid M, Seljeflot I, Weiss TW, Klemsdal TO, Hjerkinn EM, Arnesen H. (2010) Arterial stiffness is independently associated with interleukin-18 and components of the metabolic syndrome. Atherosclerosis 209(2): 337-9. |
| [44] | Nelson JH. (1985) Wheat: its processing and utilization. The American Journal of Clinical Nutrition 41(5 Suppl): 1070-76. |
| [45] | Feldman N, Norenberg C, Voet H, Manor E, Berner Y, Madar Z. (1995) Enrichment of an Israeli ethnic food with fibres and their effects on the glycaemic and insulinaemic responses in subjects with non-insulin-dependent diabetes mellitus. The British Journal of Nutrition 74(5): 681-8. |
| [46] | Latham MC. Human nutrition in the developing world (Food and Nutrition Series No. 29). Rome: Food and Agriculture Organization of the United Nations, 1997. |
| [47] | Shewry PR. (2009) Wheat. Journal of Experimental Botany 60(6): 1537. |
| [48] | Song X, Ni Z, Yao Y, Zhang Y, Sun Q. (2009) Identification of differentially expressed proteins between hybrid and parents in wheat (Triticum aestivum L.) seedling leaves. TAG Theoretical and applied genetics Theoretische und Angewandte Genetik 118(2): 213-25. |
| [49] | Gao X, Liu SW, Sun Q, Xia GM. (2010) High frequency of HMW-GS sequence variation through somatic hybridization between Agropyron elongatum and common wheat. Planta 231(2): 245-50. |
| [50] | Garg S, Pandey D, Taj G, Goel A, Kumar A. (2014) TRIPATH: A Biological Genetic and Genomic Database of Three Economically Important Fungal Pathogen of Wheat - Rust: Smut: Bunt. Bioinformation 10(7): 466-8. |
| [51] | Pomeranz Y. Wheat : chemistry and technology. 3rd ed. ed. St. Paul, Minn., USA: St. Paul, Minn., USA: American Association of Cereal Chemists, 1988. |
| [52] | de Punder K, Pruimboom L. (2013) The dietary intake of wheat and other cereal grains and their role in inflammation. Nutrients 5(3): 771-87. |
| [53] | Cecinato P, Fuccio L, Sabattini E, Laterza L, Caponi A, Azzaroli F, et al. (2014) An unusual cause of weight loss in a young Caucasian man. Common variable immunodeficiency (CVI) associated with diffuse enteral nodular lymphoid hyperplasia (NLH) and CD. Gut 63(5): 856-9. |
| [54] | Yang Y. Chinese Food Composition Table 2004: Beijing Medical University Press, 2005. |
| [55] | Prentice AM, Jebb SA. (2003) Fast foods, energy density and obesity: a possible mechanistic link. Oxford, UK2003. p. 187-94. |
| [56] | Savage JS, Marini M, Birch LL. (2008) Dietary energy density predicts women's weight change over 6 y. The American Journal of Clinical Nutrition 88(3): 677. |
| [57] | Sun Q, Spiegelman D, van Dam RM, Holmes MD, Malik VS, Willett WC, et al. (2010) White rice, brown rice, and risk of type 2 diabetes in US men and women. Archives of Internal Medicine 170(11): 961. |
| [58] | James WPT, Chunming C, Inoue S. (2002) Appropriate Asian body mass indices? (Editorial). Obesity Reviews 3(3): 139. |
| [59] | WHO/IASO/IOTF. The Asia-Pacific perspective: redefining obesity and its treatment: Health Communications Australia Pty: Melbourne. 2000. |
| [60] | Cordain L, Eaton SB, Sebastian A, Mann N, Lindeberg S, Watkins BA, et al. (2005) Origins and evolution of the Western diet: health implications for the 21st century. American Journal Of Clinical Nutrition 81(2): 341-54. |
| [61] | Grantham JP, Staub K, Rühli FJ, Henneberg M. (2014) Modern diet and metabolic variance--a recipe for disaster? Nutrition Journal 13: 01-10. |
| [62] | Speth JD. (1989) Early hominid hunting and scavenging: the role of meat as an energy source. Journal of Human Evolution 18(4): 329-43. |
| [63] | FAO and WHO. Carbohydrates in human nutrition-report of a joint FAO/WHO expert consultation. Rome: World Health Organization and Food and Agriculture Organization, 1998. |
| [64] | FAO. FAOSTAT. Available from: http://faostat3.fao.org. |
| [65] | Okubo H, Sasaki S, Murakami K, Kim MK, Takahashi Y, Hosoi Y, et al. (2007) Three major dietary patterns are all independently related to the risk of obesity among 3760 Japanese women aged 18–20 years. International Journal of Obesity 32(3): 541-9. |
| [66] | Lin H, Bermudez OI, Tucker KL. (2003) Dietary Patterns of Hispanic Elders Are Associated with Acculturation and Obesity. Journal of Nutrition 133(11): 3651-7. |
| [67] | Kim J-H, Lee JE, Jung I-K. (2012) Dietary Pattern Classifications and the Association with General Obesity and Abdominal Obesity in Korean Women. Academy of Nutrition and Dietetics 112(10): 1550-9. |
| [68] | Sichieri R. (2002) Dietary Patterns and Their Associations with Obesity in the Brazilian City of Rio de Janeiro. Obesity Research 10(1): 42-8. |
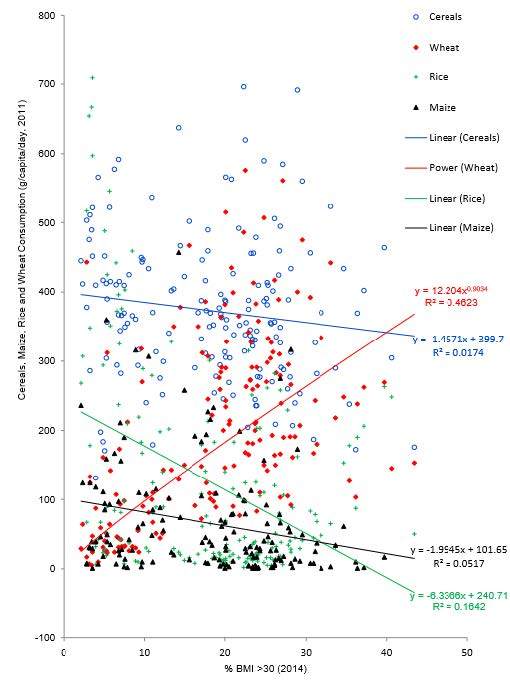
Figures(1) / Tables(4)

Wenpeng You, Maciej Henneberg. Cereal Crops Are not Created Equal: Wheat Consumption Associated with Obesity Prevalence Globally and Regionally[J]. AIMS Public Health, 2016, 3(2): 313-328. doi: 10.3934/publichealth.2016.2.313







 DownLoad:
DownLoad: