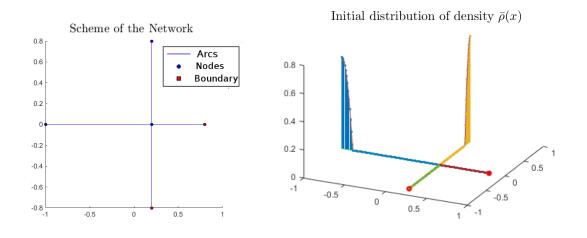
In this paper, we introduce a discrete time-finite state model for pedestrian flow on a graph in the spirit of the Hughes dynamic continuum model. The pedestrians, represented by a density function, move on the graph choosing a route to minimize the instantaneous travel cost to the destination. The density is governed by a conservation law whereas the minimization principle is described by a graph eikonal equation. We show that the discrete model is well-posed and the numerical examples reported confirm the validity of the proposed model and its applicability to describe real situations.
Citation: Fabio Camilli, Adriano Festa, Silvia Tozza. A discrete Hughes model for pedestrian flow on graphs[J]. Networks and Heterogeneous Media, 2017, 12(1): 93-112. doi: 10.3934/nhm.2017004
In this paper, we introduce a discrete time-finite state model for pedestrian flow on a graph in the spirit of the Hughes dynamic continuum model. The pedestrians, represented by a density function, move on the graph choosing a route to minimize the instantaneous travel cost to the destination. The density is governed by a conservation law whereas the minimization principle is described by a graph eikonal equation. We show that the discrete model is well-posed and the numerical examples reported confirm the validity of the proposed model and its applicability to describe real situations.

| [1] | "Jamarat: Study of Current Conditions and Means of Improvements", Hajj Research Centre, Um Al-Qura University Saudi Arabia, 1984. |
| [2] |
An efficient policy iteration algorithm for dynamic programming equations. SIAM J. Sci. Comput. (2015) 37: A181-A200. 
|
| [3] |
The one-dimensional Hughes model for pedestrian flow: Riemann-type solutions. Acta Math. Sci. (2012) 32: 259-280.
|
| [4] |
Existence results for Hughes model for pedestrian flows. J. Math. Anal. Appl. (2014) 420: 387-406.
|
| [5] |
A Dijkstra-type algorithm for dynamic games. Dyn. Games Appl. (2015) 1-4.
|
| [6] |
On the modeling of traffic and crowds: A survey of models, speculations, and perspectives. SIAM Rev. (2011) 53: 409-463.
|
| [7] |
An easy-to-use algorithm for simulating traffic flow on networks: theoretical study. Netw. Heterog. Media (2014) 9: 519-552.
|
| [8] |
A comparison among various notions of viscosity solutions for Hamilton-Jacobi equations on networks. J. Math. Anal. Appl. (2013) 407: 112-118.
|
| [9] |
Staionary mean field games systems defined on networks. SIAM J. Cont. Optim. (2016) 54: 1085-1103.
|
| [10] |
An approximation scheme for a Hamilton-Jacobi equation defined on a network. Appl. Numer. Math. (2013) 73: 33-47.
|
| [11] |
A Semi-Lagrangian scheme for a modified version of the Hughes model for pedestrian flow. Dyn. Games Appl. (2016) 1-23.
|
| [12] |
A convergent scheme for Hamilton-Jacobi equations on a junction: Application to traffic. Numer. Math. (2015) 129: 405-447.
|
| [13] |
A destination-preserving model for simulating Wardrop equilibria in traffic flow on networks. Netw. Heterog. Media (2015) 10: 857-876.
|
| [14] |
On the Hughes model for pedestrian flow: The one-dimensional case. J. Differential Equations (2011) 250: 1334-1362.
|
| [15] |
A proposed pedestrian waiting-time model for improving space-time use efficiency in stadium evacuation scenarios. Build. Environ. (2011) 46: 1774-1784.
|
| [16] | M. Garavello and B. Piccoli, "Traffic Flow on Networks" AIMS Series on Applied Mathematics, Vol. 1, American Institute of Mathematical Sciences, 2006. |
| [17] |
Revisiting Hughes dynamic continuum model for pedestrian flow and the development of an efficient solution algorithm. Transportat. Res. B-Meth. (2009) 43: 127-141.
|
| [18] |
The flow of large crowds of pedestrians. Math. Comput. Simulat. (2000) 53: 367-370.
|
| [19] |
A continuum theory for the flow of pedestrians. Transport. Res. B-Meth. (2002) 36: 507-535.
|
| [20] |
The flow of human crowds. Annu. rev. fluid mech. (2003) 35: 169-182.
|
| [21] |
Viscosity solutions for junctions: Well posedness and stability. Rend. Lincei Mat. Appl. (2016) 27: 535-545.
|
| [22] | Nonlinear elliptic partial differential equations and p-harmonic functions on graphs. Differ. Integral Equ. (2015) 28: 79-102. |
| [23] |
On the convergence of policy iteration in stationary dynamic programming. Math. Oper. Res. (1979) 4: 60-69.
|
| [24] |
Convergence of a difference scheme for conservation laws with a discontinuous flux. SIAM J. Numer. Anal. (2000) 38: 681-698.
|
| [25] |
Continuum crowds. ACM Trans. Graph. (2006) 25: 1160-1168.
|







Figures(8)
Fabio Camilli, Adriano Festa, Silvia Tozza. A discrete Hughes model for pedestrian flow on graphs[J]. Networks and Heterogeneous Media, 2017, 12(1): 93-112. doi: 10.3934/nhm.2017004







 DownLoad:
DownLoad: