Citation: Paola F. Antonietti, Ilario Mazzieri, Laura Melas, Roberto Paolucci, Alfio Quarteroni, Chiara Smerzini, Marco Stupazzini. Three-dimensional physics-based earthquake ground motion simulations for seismic risk assessment in densely populated urban areas[J]. Mathematics in Engineering, 2021, 3(2): 1-31. doi: 10.3934/mine.2021012

| [1] | Smolka A, Allmann A, Hollnack D, et al. (2004) The principle of risk partnership and the role of insurance in risk mitigation, In: Proceedings of the 13th World Conference on Earthquake Engineering, 2020. |
| [2] | Erdik M (2017) Earthquake risk assessment. B Earthq Eng 15: 5055-5092. |
| [3] | Douglas J, Aochi H (2008) A survey of techniques for predicting earthquake ground motions for engineering purposes. Surv Geophys 29: 187. |
| [4] | Douglas J, Edwards B (2016) Recent and future developments in earthquake ground motion estimation. Earth-Sci Rev 160: 203-219. |
| [5] | Peruš I, Fajfar P (2009) How reliable are the ground motion prediction equations, In: Proceedings of the 20th International Conference on Structural Mechanics in Reactor Technology (SMiRT 20), Espoo, 1662. |
| [6] | Al Atik L, Abrahamson N, Bommer J, et al. (2010) The variability of ground-motion prediction models and its components. Seismol Res Lett 81: 794-801. |
| [7] | Jayaram N, Baker J (2009) Correlation model for spatially distributed ground-motion intensities. Earthq Eng Struct D 38: 1687-1708. |
| [8] | Park J, Bazzurro P, Baker J (2007) Modeling spatial correlation of ground motion intensity measures for regional seismic hazard and portfolio loss estimation, In: Applications of Statistics and Probability in Civil Engineering - Proceedings of the 10th International Conference on Applications of Statistics and Probability, ICASP10. |
| [9] | Weatherill G, Silva V, Crowley H, et al. (2015) Exploring the impact of spatial correlations and uncertainties for portfolio analysis in probabilistic seismic loss estimation. B Earthq Eng 13: 957-981. |
| [10] | Antonietti PF, Dal Santo N, Mazzieri I, et al. (2018) A high-order discontinuous Galerkin approximation to ordinary differential equations with applications to elastodynamics. IMA J Numer Anal 38: 1709-1734. |
| [11] | Bradley B (2018) On-going challenges in physics-based ground motion prediction and insights from the 2010-2011 Canterbury and 2016 Kaikoura, New Zealand earthquakes. Soil Dyn Earthq Eng 124: 354-364. |
| [12] | Graves RW (1996) Simulating seismic wave propagation in 3D elastic media using staggered-grid finite differences. B Seismol Soc Am 86: 1091-1106. |
| [13] | Lysmer J, Drake LA (1972) A finite element method for seismology, In: Seismology: Surface Waves and Earth Oscillations, Academic Press Inc., 181-216. |
| [14] | Faccioli E, Maggio F, Paolucci R, et al. (1997) 2D and 3D elastic wave propagation by a pseudospectral domain decomposition method. J Seismol 1: 237-251. |
| [15] | Komatitsch D, Vilotte JP (1998) The spectral element method: An efficient tool to simulate the seismic response of 2D and 3D geological structures. B Seismol Soc Am 88: 368-392. |
| [16] | Villani M, Faccioli E, Ordaz M, et al. (2014) High resolution seismic hazard analysis in a complex geological configuration: The case of the Sulmona basin in Central Italy. Earthq Spectra 30: 1801-1824. |
| [17] | Paolucci R, Mazzieri I, Smerzini C (2015) Anatomy of strong ground motion: Near-source records and three-dimensional physics-based numerical simulations of the Mw 6.0 2012 May 29 Po plain earthquake, Italy. Geophys J Int 203: 2001-2020. |
| [18] | Paolucci R, Evangelista L, Mazzieri I, et al. (2016) The 3D numerical simulation of near-source ground motion during the Marsica earthquake, Central Italy, 100 years later. Soil Dyn Earthq Eng 91: 39-52. |
| [19] | Antonietti PF, Mazzieri I, Quarteroni A, et al. (2012) Non-conforming high order approximations of the elastodynamics equation. Comput Method Appl M 209: 212-238. |
| [20] | Käser M, Dumbser M (2006) An arbitrary high-order discontinuous Galerkin method for elastic waves on unstructured meshes-I. The two-dimensional isotropic case with external source terms. Geophys J Int 166: 855-877. |
| [21] | Antonietti PF, Ayuso de Dios B, Mazzieri I, et al. (2016) Stability analysis of discontinuous Galerkin approximations to the elastodynamics problem. J Sci Comput 68: 143-170. |
| [22] | Mazzieri I, Stupazzini M, Guidotti R, et al. (2013) SPEED: SPectral Elements in Elastodynamics with Discontinuous Galerkin: A non-conforming approach for 3D multi-scale problems. Int J Numer Meth Eng 95: 991-1010. |
| [23] | Antonietti PF, Mazzieri I (2018) High-order Discontinuous Galerkin methods for the elastodynamics equation on polygonal and polyhedral meshes. Comput Method Appl M 342: 414-437. |
| [24] | Ferroni A, Antonietti PF, Mazzieri I, et al. (2017) Dispersion-dissipation analysis of 3-D continuous and discontinuous spectral element methods for the elastodynamics equation. Geophys J Int 211: 1554-1574. |
| [25] | Graves R, Jordan T, Callaghan S, et al. (2011) CyberShake: A physics-based seismic hazard model for Southern California. Pure Appl Geophys 168: 367-381. |
| [26] | Paolucci R, Infantino M, Mazzieri I, et al. (2018) 3D physics-based numerical simulations: Advantages and current limitations of a new frontier to earthquake ground motion prediction. The Istanbul case study, In: Recent Advances in Earthquake Engineering in Europe: 16th European Conference on Earthquake Engineering-Thessaloniki 2018, Springer, 203-223. |
| [27] | Infantino M, Mazzieri I, Özcebe A, et al. (2020) 3D physics-based numerical simulations of ground motion in Istanbul from earthquakes along the Marmara segment of the North Anatolian Fault. Bull seism Soc Am. |
| [28] | Porter K, Jones L, Cox D, et al. (2011) The ShakeOut scenario: A hypothetical Mw7.8 earthquake on the Southern San Andreas fault. Earthq Spectra 27: 239-261. |
| [29] | Smerzini C, Pitilakis K (2018) Seismic risk assessment at urban scale from 3D physics-based numerical modeling: The case of Thessaloniki. B Earthq Eng 16: 2609-2631. |
| [30] | Detweiler S, Wein A (2017) The HayWired earthquake scenario-Earthquake hazards. Scientific Investigations Report 2017-5013(A-H). U.S Geological Survey. |
| [31] | Detweiler S, Wein A (2018) The HayWired earthquake scenario-Engineering implications. Scientific Investigations Report 2017-5013(I-Q). U.S Geological Survey. |
| [32] | Evangelista L, del Gaudio S, Smerzini C, et al. (2017) Physics-based seismic input for engineering applications: A case study in the Aterno river valley, Central Italy. B Earthq Eng 15: 2645-2671. |
| [33] | Guidotti R, Stupazzini M, Smerzini C, et al. (2011) Numerical study on the role of basin geometry and kinematic seismic source in 3D ground motion simulation of the 22 February 2011 Mw 6.2 Christchurch earthquake. Seismol Res Lett 82: 767-782. |
| [34] | Smerzini C, Pitilakis K, Hashemi K (2017) Evaluation of earthquake ground motion and site effects in the Thessaloniki urban area by 3D finite-fault numerical simulations. B Earthq Eng 15: 787-812. |
| [35] | Stacey R (1988) Improved transparent boundary formulations for the elastic-wave equation. B Seismol Soc Am 78: 2089-2097. |
| [36] | Antonietti PF, Ferroni A, Mazzieri I, et al. (2018) Numerical modeling of seismic waves by discontinuous spectral element methods. ESAIM ProcS 61: 1-37. |
| [37] | Aki K, Richards PG (2002) Quantitive Seismology: Theory and Methods. San Francisco: Freeman. |
| [38] | Arnold DN, Brezzi F, Cockburn B, et al. (2002) Unified analysis of discontinuous Galerkin methods for elliptic problems. SIAM J Numer Anal 39: 1749-1779. |
| [39] | Arnold DN (1982) An interior penalty finite element method with discontinuous elements. SIAM J Numer Anal 19: 742-760. |
| [40] | Epshteyn Y, Rivière B (2007) Estimation of penalty parameters for symmetric interior penalty Galerkin methods. J Comput Appl Math 206: 843-872. |
| [41] | Rivière B, Wheeler MF (2003) Discontinuous finite element methods for acoustic and elastic wave problems, In: Current trends in scientific computing (Xi'an, 2002), Providence: Amer. Math. Soc., 271-282. |
| [42] | Rivière B, Shaw S, Whiteman JR (2007) Discontinuous Galerkin finite element methods for dynamic linear solid viscoelasticity problems. Numer Meth Part D E 23: 1149-1166. |
| [43] | Canuto C, Hussaini MY, Quarteroni A, et al. (2006) Spectral Methods - Fundamentals in Single Domains, Berlin: Springer-Verlag. |
| [44] | Quarteroni A, Valli A (1994) Numerical Approximation of Partial Differential Equations, Berlin: Springer-Verlag. |
| [45] | Canuto C, Hussaini MY, Quarteroni A, et al. (2007) Spectral methods - Evolution to complex geometries and applications to fluid dynamics. Berlin: Springer. |
| [46] | di Prisco C, Stupazzini M, Zambelli C (2007) Nonlinear SEM numerical analyses of dry dense sand specimens under rapid and dynamic loading. Int J Numer Anal Met 31: 757-788. |
| [47] | Stupazzini M, Paolucci R, Igel H (2009) Near-fault earthquake ground-motion simulation in the Grenoble valley by a high-performance spectral element code. B Seismol Soc Am 99: 286-301. |
| [48] | Kramer SL (1996) Earthquake Geotechnical Engineering, Pearson Education India. |
| [49] | Luco N, Cornell C (2007) Structure-specific scalar intensity measures for near-source and ordinary earthquake ground motions. Earthq Spectra 23: 357-392. |
| [50] | Housner GW (1952) Spectrum intensities of strong-motion earthquakes. Earthq Eng Res Inst 21-36. |
| [51] | Mai C, Konakli K, Sudret B (2017) Seismic fragility curves for structures using non-parametric representations. Front Struct Civ Eng 11: 169-186. |
| [52] | Singhal A, Kiremidjian AS (1996) Method for probabilistic evaluation of seismic structural damage. J Struct Eng 122: 1459-1467. |
| [53] | Shinozuka M, Feng MQ, Lee J, et al. (2000) Statistical analysis of fragility curves. J Eng Mech 126: 1224-1231. |
| [54] | Ellingwood BR (2001) Earthquake risk assessment of building structures. Reliab Eng Syst Safe 74: 251-262. |
| [55] | Porter K, Kennedy R, Bachman R (2007) Creating fragility functions for performance-based earthquake engineering. Earthq Spectra 23: 471-489. |
| [56] | Seyedi D, Gehl P, Douglas J, et al. (2010) Development of seismic fragility surfaces for reinforced concrete buildings by means of nonlinear time-history analysis. Earthq Eng Struct D 39: 91-108. |
| [57] | Zentner I (2010) Numerical computation of fragility curves for NPP equipment. Nucl Eng Des 240: 1614-1621. |
| [58] | Gencturk B, Elnashai AS, Song J (2008) Fragility relationships for populations of woodframe structures based on inelastic response. J Earthq Eng 12: 119-128. |
| [59] | Jeong SH, Mwafy AM, Elnashai AS (2012) Probabilistic seismic performance assessment of codecompliant multi-story RC buildings. Eng Struct 34: 527-537. |
| [60] | Banerjee S, Shinozuka M (2008) Mechanistic quantification of RC bridge damage states under earthquake through fragility analysis. Probabilist Eng Mech 23: 12-22. |
| [61] | Wu F, Wang M, Yang XY (2013) Building seismic vulnerability study for China high rises. Appl Mech Mater 353: 2301-2304. |
| [62] | Gu G, Lin T, Shi Z (1983) Catalogue of Earthquakes in China (1831AD-1969BC). Beijing: Science Press. |
| [63] | Ding Z, Romanelli F, Chen Y, et al. (2004) Realistic modeling of seismic wave ground motion in Beijing city. Pure Appl Geophys 161: 1093-1106. |
| [64] | Gao M, Yu Y, Zhang X, et al. (2004) Three-dimensional finite-difference modeling of ground motions in Beijing form a Mw 7 scenario earthquake, In: Proceedings of the 13th World Conference on Earthquake Engineering, 581. |
| [65] | Xiong C, Lu X, Huang J, et al. (2019) Multi-LOD seismic-damage simulation of urban buildings and case study in Beijing CBD. B Earthq Eng 17: 2037-2057. |
| [66] | Xu Z, Lu X, Zeng X, et al. (2019) Seismic loss assessment for buildings with various-LOD BIM data. Adv Eng Inform 39: 112-126. |
| [67] | Lu X, Zeng X, Xu Z, et al. (2019) Improving the accuracy of near real-time seismic loss estimation using post-earthquake remote sensing images. Earthq Spectra 34: 1219-1245. |
| [68] | Allen TI, Wald DJ (2009) On the use of high-resolution topographic data as a proxy for seismic site conditions (VS30). B Seismol Soc Am 99: 935-943. |
| [69] | Wells DL, Coppersmith KJ (1994) New empirical relationships among magnitude, rupture length, rupture width, rupture area, and surface displacement. B Seismol Soc Am 84: 974-1002. |
| [70] | Causse M, Cotton F, Cornou C, et al. (2008) Calibrating median and uncertainty estimates for a practical use of empirical Green's functions technique. B Seismol Soc Am 98: 344-353. |
| [71] | Schmedes J, Archuleta RJ, Lavallée D (2012) A kinematic rupture model generator incorporating spatial interdependency of earthquake source parameters. Geophys J Int 192: 1116-1131. |
| [72] | Cauzzi C, Faccioli E, Vanini M, et al. (2015) Updated predictive equations for broadband (0.01-10 s) horizontal response spectra and peak ground motions, based on a global dataset of digital acceleration records. B Earthq Eng 13: 1587-1612. |
| [73] | Moehle J, Bozorgnia Y, Jayaram N, et al. (2011) Case studies of the seismic performance of tall buildings designed by alternative means. Pacific Earthquake Engineering Research Center College of Engineering University of California, Berkeley PEER Report 5. |
| [74] | Kazantzi A, Vamvatsikos D, Porter K, et al. (2014) Analytical vulnerability assessment of modern highrise RC moment-resisting frame buildings in the Western USA for the Global Earthquake Model, In: Proceedings of the 2nd European Conference on Earthquake Engineering and Seismology. |
| [75] | Council BSS (1997) NEHRP guidelines for the seismic rehabilitation of buildings. FEMA-273, Federal Emergency Management Agency, Washington, DC. |
| [76] | Xu P, Xiao C, Li J (2014) Research on relationship between natural vibration periods and structural heights for high-rise buildings and its reference range in China. Int J High-rise Buildings 3: 49-64. |
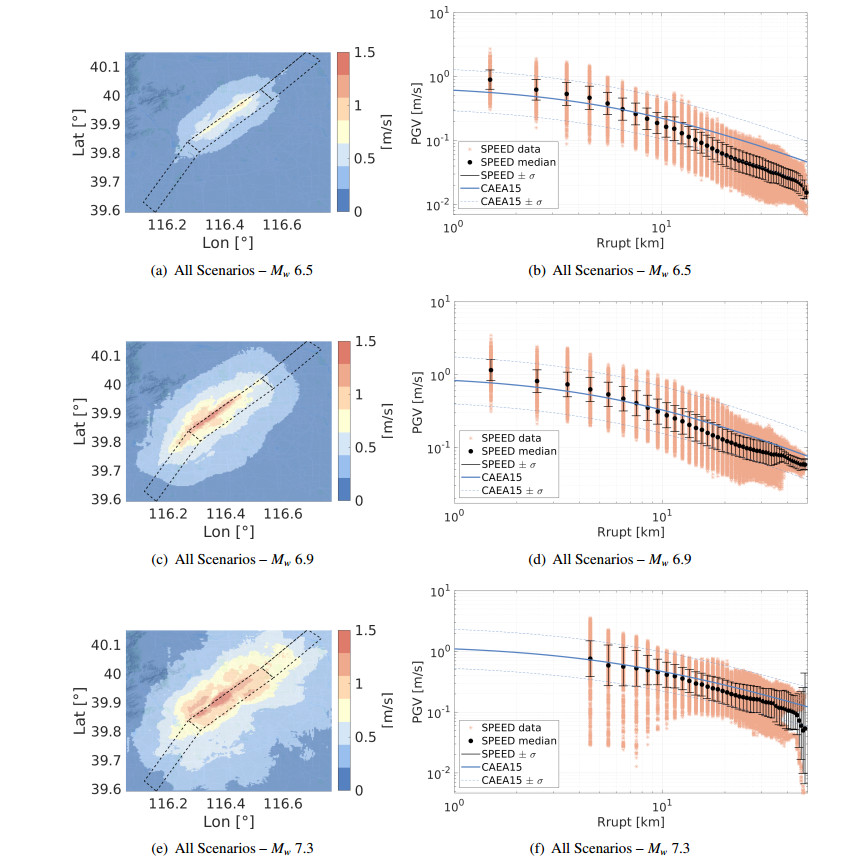
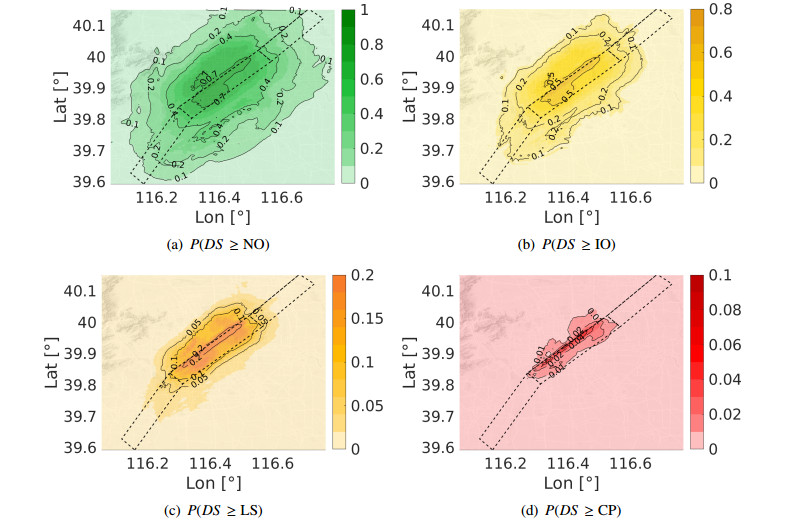
Figures(10) / Tables(8)

Paola F. Antonietti, Ilario Mazzieri, Laura Melas, Roberto Paolucci, Alfio Quarteroni, Chiara Smerzini, Marco Stupazzini. Three-dimensional physics-based earthquake ground motion simulations for seismic risk assessment in densely populated urban areas[J]. Mathematics in Engineering, 2021, 3(2): 1-31. doi: 10.3934/mine.2021012







 DownLoad:
DownLoad: