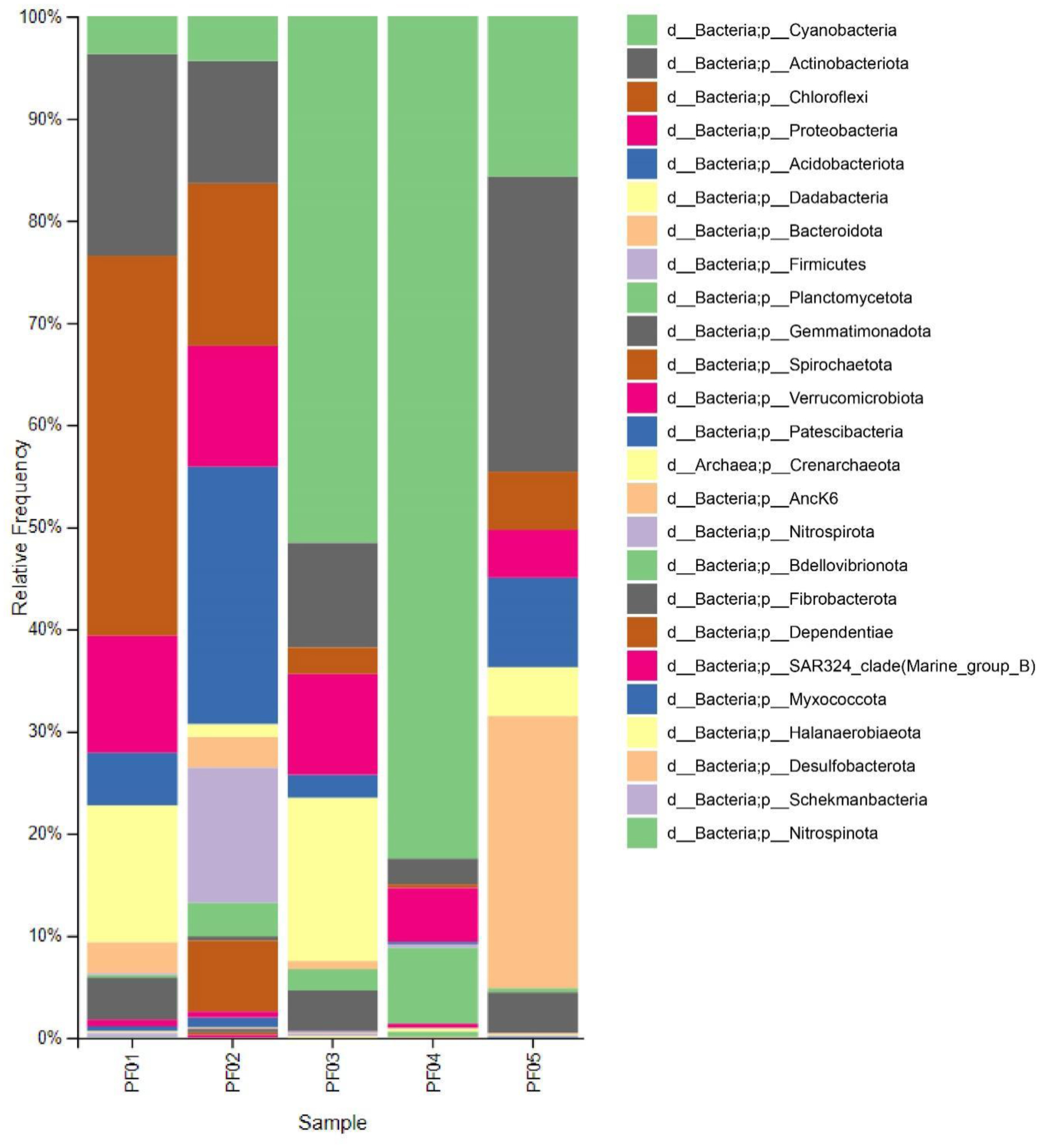
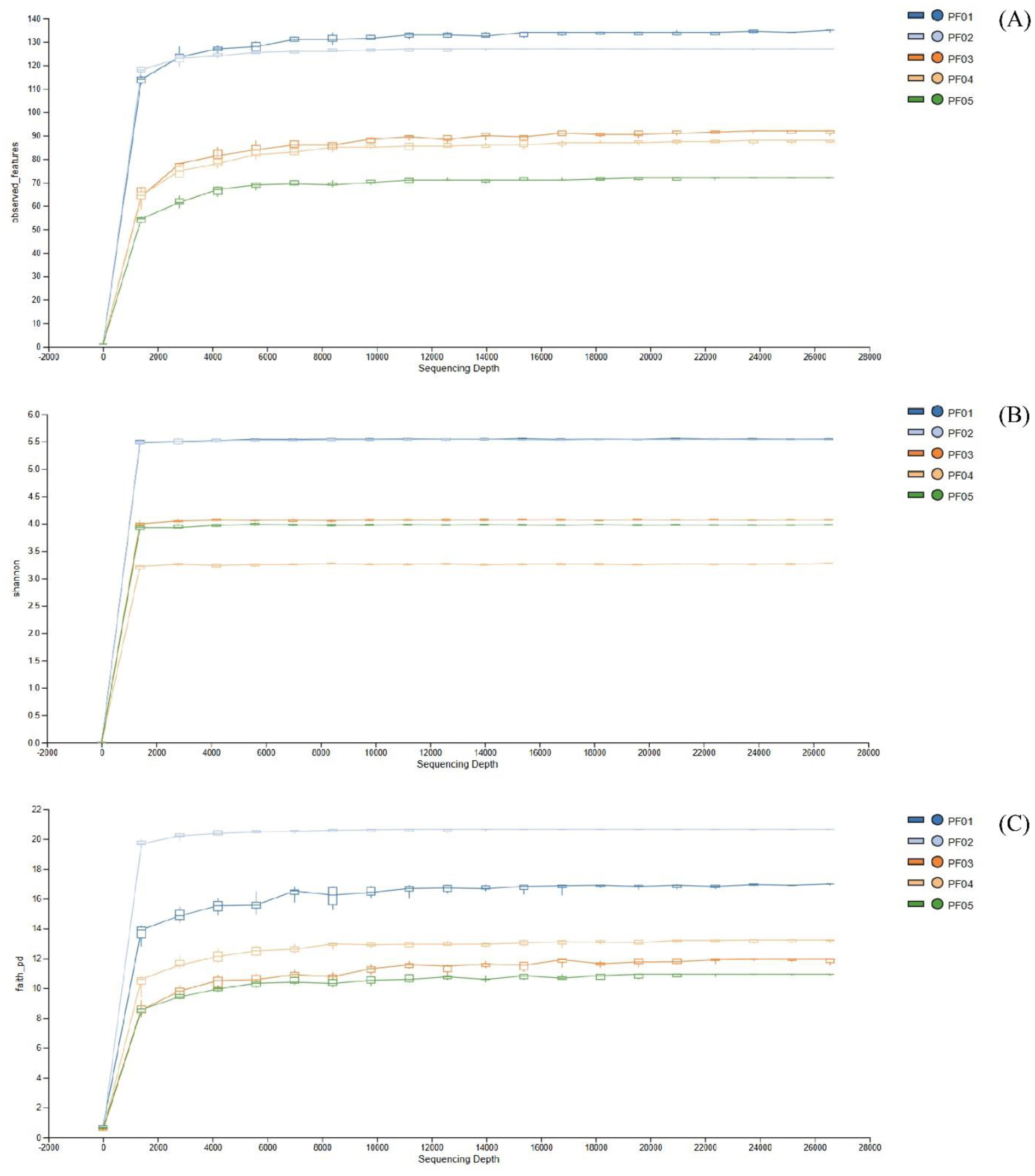
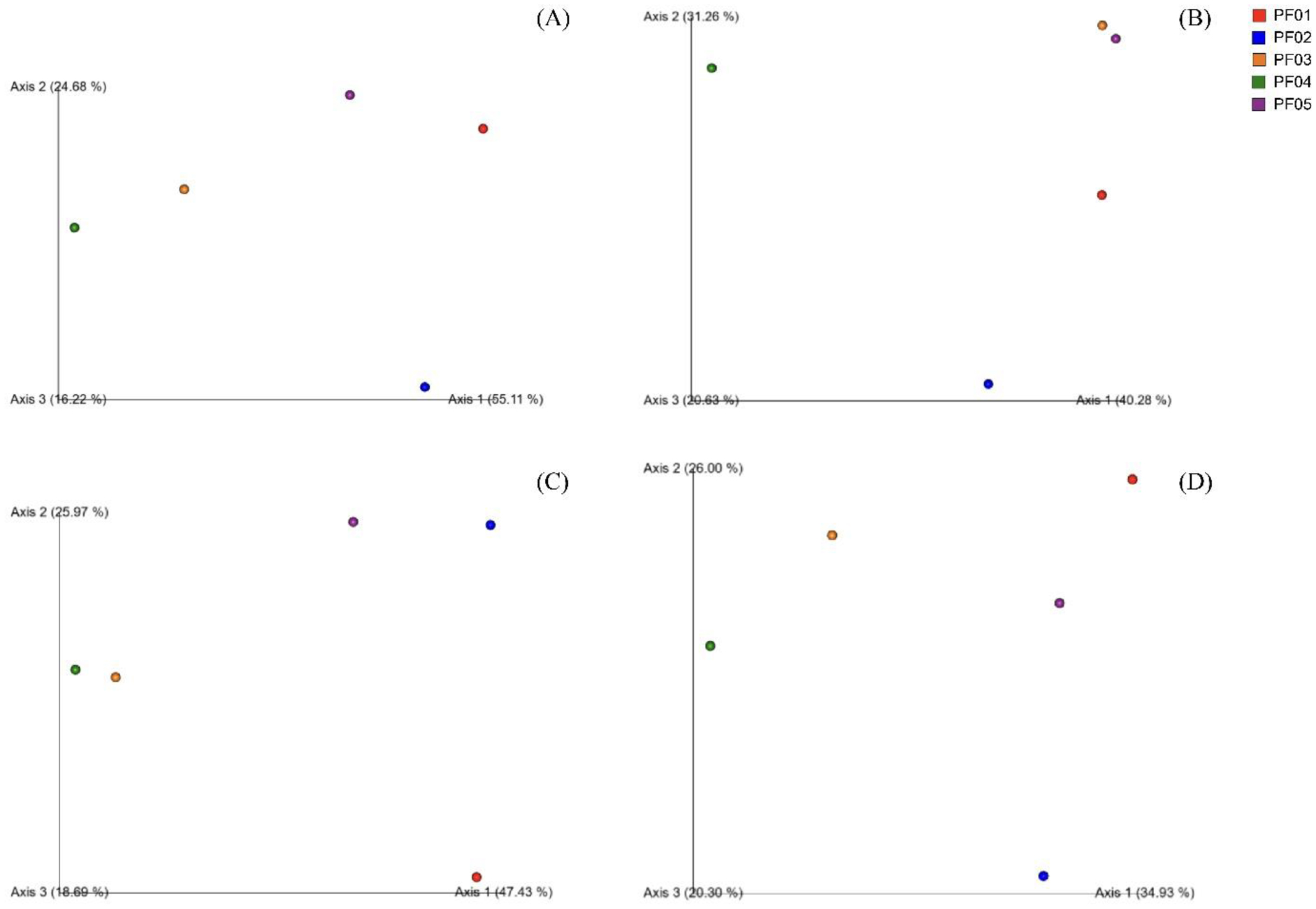
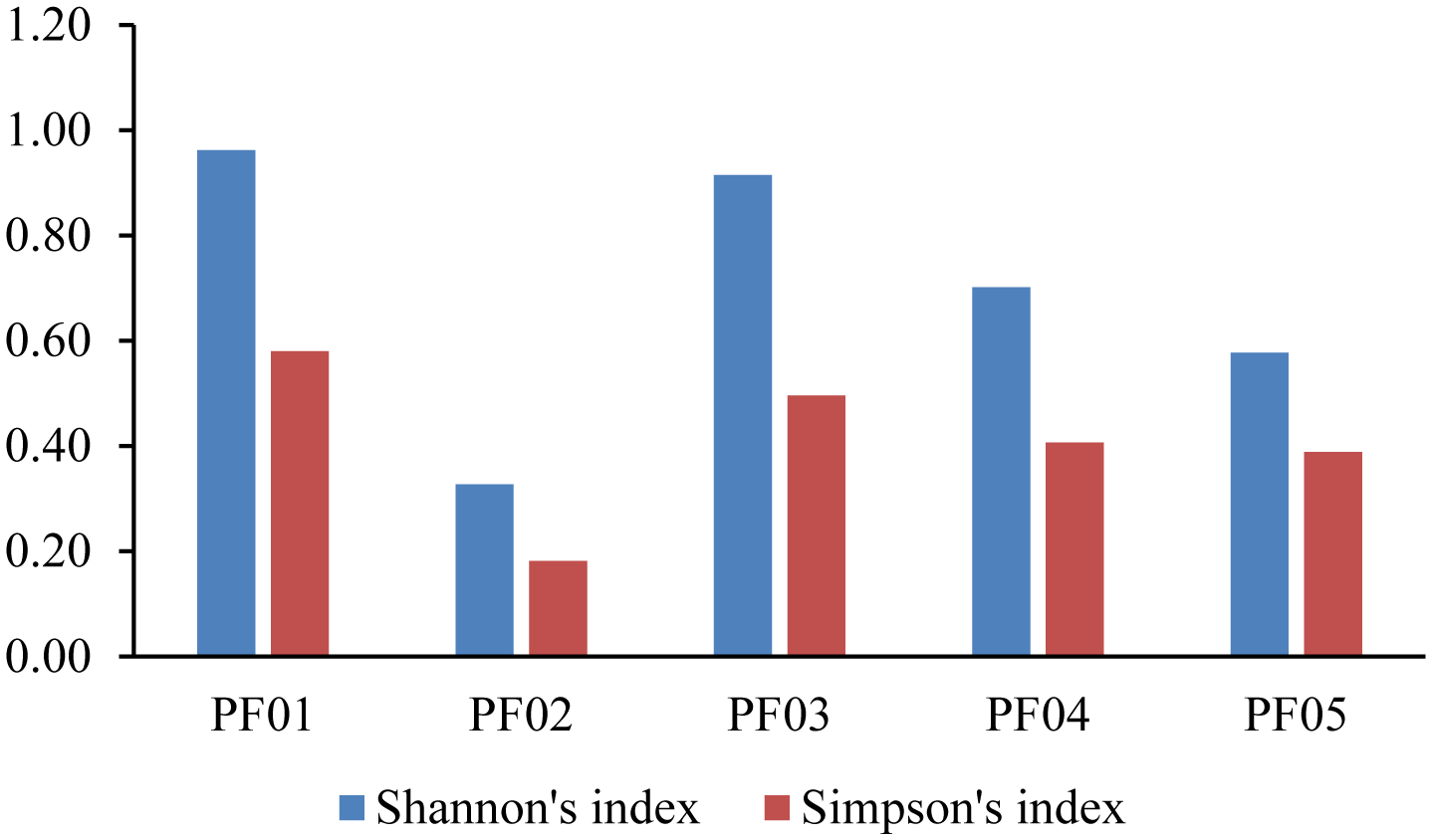
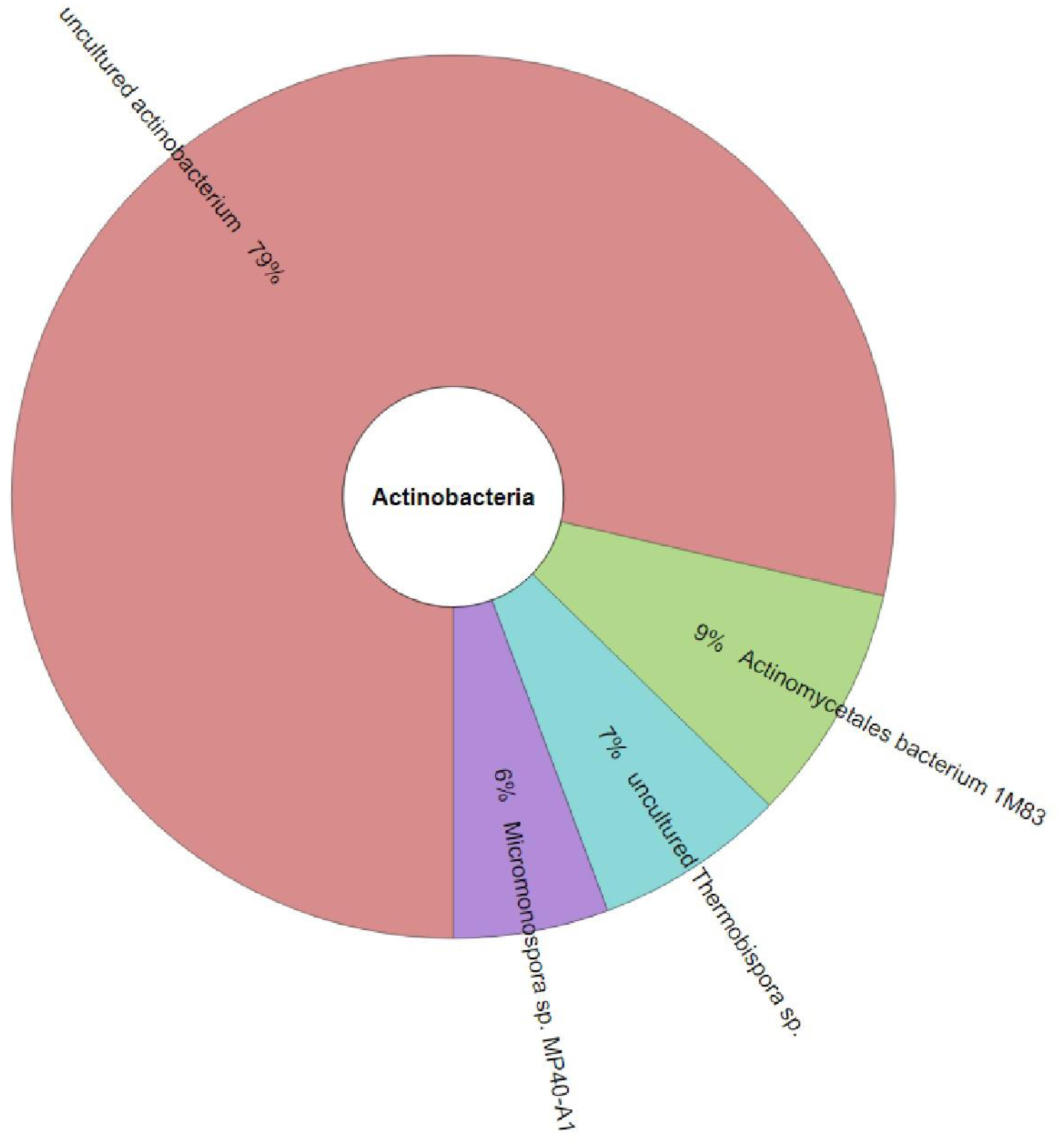
Marine sponges are well-known for their production of bioactive compounds, many of which are synthesized by their associated symbiotic microorganisms. Among these, Actinomycetes are of particular interest due to their ability to produce secondary metabolites with antimicrobial and antitumor activities. We aimed to investigate the bacterial microbiome of tropical marine sponges, with an emphasis on the diversity and distribution of Actinomycetes, employing both culture-dependent and culture-independent approaches. Five sponge samples (PF01–PF05) were collected from Sichang Island, Chonburi Province, Thailand. The bacterial communities were analyzed using 16S rRNA gene sequencing and bioinformatics tools, revealing a significant microbial diversity dominated by Cyanobacteria, Actinomycetota, and Chloroflexi. Notably, PF01 (Penares nux) exhibited the highest microbial diversity, while PF05 (Cacospongia sp.) had the lowest. Actinomycetes, particularly the genus Micromonospora, were successfully isolated from all samples, with PF03 (Ircinia mutans) yielding the highest number of strains. Culture-independent analysis identified a greater proportion of unculturable Actinomycetes compared to those isolated through traditional methods, underscoring the limitations of culture-dependent techniques. This study enhances our understanding of sponge-associated microbial diversity and highlights the potential for isolating Actinomycetes from these sponges for novel drug discovery and other bioprospective applications.
Citation: Trinset Weeraphan, Chollabuppha Chou, Naphatson Chanthathamrongsiri, Thanchanok Sirirak, Sumaitt Putchakarn, Supakarn Chamni, Wongsakorn Phongsopitanun. Microbiome profiling and Actinomycetes isolation from tropical marine sponges[J]. AIMS Microbiology, 2025, 11(1): 182-205. doi: 10.3934/microbiol.2025010
Marine sponges are well-known for their production of bioactive compounds, many of which are synthesized by their associated symbiotic microorganisms. Among these, Actinomycetes are of particular interest due to their ability to produce secondary metabolites with antimicrobial and antitumor activities. We aimed to investigate the bacterial microbiome of tropical marine sponges, with an emphasis on the diversity and distribution of Actinomycetes, employing both culture-dependent and culture-independent approaches. Five sponge samples (PF01–PF05) were collected from Sichang Island, Chonburi Province, Thailand. The bacterial communities were analyzed using 16S rRNA gene sequencing and bioinformatics tools, revealing a significant microbial diversity dominated by Cyanobacteria, Actinomycetota, and Chloroflexi. Notably, PF01 (Penares nux) exhibited the highest microbial diversity, while PF05 (Cacospongia sp.) had the lowest. Actinomycetes, particularly the genus Micromonospora, were successfully isolated from all samples, with PF03 (Ircinia mutans) yielding the highest number of strains. Culture-independent analysis identified a greater proportion of unculturable Actinomycetes compared to those isolated through traditional methods, underscoring the limitations of culture-dependent techniques. This study enhances our understanding of sponge-associated microbial diversity and highlights the potential for isolating Actinomycetes from these sponges for novel drug discovery and other bioprospective applications.

| [1] |
Van Soest RW, Boury-Esnault N, Vacelet J, et al. (2012) Global diversity of sponges (Porifera). PLoS One 7: e35105. https://doi.org/10.1371/journal.pone.0035105 
|
| [2] |
Lavrov DV, Diaz MC, Maldonado M, et al. (2023) Phylomitogenomics bolsters the high-level classification of Demospongiae (phylum Porifera). PLoS One 18: e0287281. https://doi.org/10.1371/journal.pone.0287281
|
| [3] |
Jahn MT, Lachnit T, Markert SM, et al. (2021) Lifestyle of sponge symbiont phages by host prediction and correlative microscopy. ISME J 15: 2001-2011. https://doi.org/10.1038/s41396-021-00900-6
|
| [4] |
Thacker RW, Freeman CJ (2012) Sponge-microbe symbioses: recent advances and new directions. Adv Mar Biol 62: 57-111. https://doi.org/10.1016/B978-0-12-394283-8.00002-3
|
| [5] |
Laport MS, Santos OC, Muricy G (2009) Marine sponges: potential sources of new antimicrobial drugs. Curr Pharm Biotechnol 10: 86-105. https://doi.org/10.2174/138920109787048625
|
| [6] |
Varijakzhan D, Loh JY, Yap WS, et al. (2021) Bioactive compounds from marine sponges: Fundamentals and applications. Mar Drugs 19. https://doi.org/10.3390/md19050246
|
| [7] |
Wakimoto T (2023) Biosynthesis of bioactive natural products derived from Theonellidae family marine sponges. Chem Pharm Bull (Tokyo) 71: 1-8. https://doi.org/10.1248/cpb.c22-00715
|
| [8] |
Di Cesare Mannelli L, Palma Esposito F, Sangiovanni E, et al. (2021) Pharmacological activities of extracts and compounds isolated from mediterranean sponge sources. Pharmaceuticals (Basel) 14. https://doi.org/10.3390/ph14121329
|
| [9] | Dinarvand M, Spain M (2021) Identification of bioactive compounds from marine natural products and exploration of structure-activity relationships (SAR). Antibiotics (Basel) 10. https://doi.org/10.3390/antibiotics10030337 |
| [10] |
Bibi F, Faheem M, Azhar EI, et al. (2017) Bacteria from marine sponges: A source of new drugs. Curr Drug Metab 18: 11-15. https://doi.org/10.2174/1389200217666161013090610
|
| [11] |
Taylor MW, Radax R, Steger D, et al. (2007) Sponge-associated microorganisms: evolution, ecology, and biotechnological potential. Microbiol Mol Biol Rev 71: 295-347. https://doi.org/10.1128/MMBR.00040-06
|
| [12] |
Brinkmann CM, Marker A, Kurtböke Dİ (2017) An overview on marine sponge-symbiotic bacteria as unexhausted sources for natural product discovery. Diversity 9: 40. https://doi.org/10.3390/d9040040
|
| [13] |
Skariyachan S, A GR, Patil MR, et al. (2014) Antimicrobial potential of metabolites extracted from bacterial symbionts associated with marine sponges in coastal area of Gulf of Mannar Biosphere, India. Lett Appl Microbiol 58: 231-241. https://doi.org/10.1111/lam.12178
|
| [14] |
Bibi F, Yasir M, Al-Sofyani A, et al. (2020) Antimicrobial activity of bacteria from marine sponge Suberea mollis and bioactive metabolites of Vibrio sp. EA348. Saudi J Biol Sci 27: 1139-1147. https://doi.org/10.1016/j.sjbs.2020.02.002
|
| [15] |
Jagannathan SV, Manemann EM, Rowe SE, et al. (2021) Marine actinomycetes, new sources of biotechnological products. Mar Drugs 19. https://doi.org/10.3390/md19070365
|
| [16] |
Lu S, Wang J, Sheng R, et al. (2020) Novel bioactive polyketides isolated from marine actinomycetes: An update review from 2013 to 2019. Chem Biodivers 17: e2000562. https://doi.org/10.1002/cbdv.202000562
|
| [17] |
De Rop AS, Rombaut J, Willems T, et al. (2021) Novel alkaloids from marine Actinobacteria: discovery and characterization. Mar Drugs 20. https://doi.org/10.3390/md20010006
|
| [18] | Chen J, Xu L, Zhou Y, et al. (2021) Natural products from Actinomycetes associated with marine organisms. Mar Drugs 19. https://doi.org/10.3390/md19110629 |
| [19] |
Yang C, Qian R, Xu Y, et al. (2019) Marine Actinomycetes-derived natural products. Curr Top Med Chem 19: 2868-2918. https://doi.org/10.2174/1568026619666191114102359
|
| [20] | Gajdács M, Urbán E (2020) The pathogenic role of Actinomyces spp. and related organisms in genitourinary infections: Discoveries in the new, modern diagnostic era. Antibiotics (Basel) 9. https://doi.org/10.3390/antibiotics9080524 |
| [21] |
Siavashifar M, Rezaei F, Motallebirad T, et al. (2021) Species diversity and molecular analysis of opportunistic Mycobacterium, Nocardia and Rhodococcus isolated from the hospital environment in a developing country, a potential resources for nosocomial infection. Genes Environ 43: 2. https://doi.org/10.1186/s41021-021-00173-7
|
| [22] | De Simeis D, Serra S (2021) Actinomycetes: A never-ending source of bioactive compounds-an overview on antibiotics production. Antibiotics (Basel) 10. https://doi.org/10.3390/antibiotics10050483 |
| [23] |
Alam K, Mazumder A, Sikdar S, et al. (2022) Streptomyces: The biofactory of secondary metabolites. Front Microbiol 13: 968053. https://doi.org/10.3389/fmicb.2022.968053
|
| [24] | Waters M, Tadi P (2024) Streptomycin. StatPearls . Treasure Island (FL): StatPearls Publishing Copyright © 2024, StatPearls Publishing LLC. |
| [25] |
Quinn GA, Banat AM, Abdelhameed AM, et al. (2020) Streptomyces from traditional medicine: sources of new innovations in antibiotic discovery. J Med Microbiol 69: 1040-1048. https://doi.org/10.1099/jmm.0.001232
|
| [26] |
Procópio RE, Silva IR, Martins MK, et al. (2012) Antibiotics produced by Streptomyces. Braz J Infect Dis 16: 466-471. https://doi.org/10.1016/j.bjid.2012.08.014
|
| [27] |
Ngamcharungchit C, Chaimusik N, Panbangred W, et al. (2023) Bioactive metabolites from terrestrial and marine Actinomycetes. Molecules 28. https://doi.org/10.3390/molecules28155915
|
| [28] |
Selim MSM, Abdelhamid SA, Mohamed SS (2021) Secondary metabolites and biodiversity of actinomycetes. J Genet Eng Biotechnol 19: 72. https://doi.org/10.1186/s43141-021-00156-9
|
| [29] |
Wang C, Lu Y, Cao S (2020) Antimicrobial compounds from marine actinomycetes. Arch Pharm Res 43: 677-704. https://doi.org/10.1007/s12272-020-01251-0
|
| [30] |
Olano C, Méndez C, Salas JA (2009) Antitumor compounds from marine actinomycetes. Mar Drugs 7: 210-248. https://doi.org/10.3390/md7020210
|
| [31] |
Ogawara H (2016) Self-resistance in Streptomyces, with special reference to β-lactam antibiotics. Molecules 21. https://doi.org/10.3390/molecules21050605
|
| [32] |
Supong K, Sripreechasak P, Phongsopitanun W, et al. (2019) Antimicrobial substances from the rare actinomycete Nonomuraea rhodomycinica NR4-ASC07(T). Nat Prod Res 33: 2285-2291. https://doi.org/10.1080/14786419.2018.1440223
|
| [33] |
Al-Fadhli AA, Threadgill MD, Mohammed F, et al. (2022) Macrolides from rare actinomycetes: Structures and bioactivities. Int J Antimicrob Agents 59: 106523. https://doi.org/10.1016/j.ijantimicag.2022.106523
|
| [34] |
Parra J, Beaton A, Seipke RF, et al. (2023) Antibiotics from rare actinomycetes, beyond the genus Streptomyces. Curr Opin Microbiol 76: 102385. https://doi.org/10.1016/j.mib.2023.102385
|
| [35] |
Tiwari K, Gupta RK (2012) Rare actinomycetes: a potential storehouse for novel antibiotics. Crit Rev Biotechnol 32: 108-132. https://doi.org/10.3109/07388551.2011.562482
|
| [36] | Sapkota A, Thapa A, Budhathoki A, et al. (2020) Isolation, characterization, and screening of antimicrobial-producing actinomycetes from soil samples. Int J Microbiol 2020: 2716584. https://doi.org/10.1155/2020/2716584 |
| [37] |
Gebreyohannes G, Moges F, Sahile S, et al. (2013) Isolation and characterization of potential antibiotic producing actinomycetes from water and sediments of Lake Tana, Ethiopia. Asian Pac J Trop Biomed 3: 426-435. https://doi.org/10.1016/S2221-1691(13)60092-1
|
| [38] | Mondal H, Thomas J (2022) Isolation and characterization of a novel Actinomycete isolated from marine sediments and its antibacterial activity against fish pathogens. Antibiotics (Basel) 11. https://doi.org/10.3390/antibiotics11111546 |
| [39] |
Fahmy NM, Abdel-Tawab AM (2021) Isolation and characterization of marine sponge-associated Streptomyces sp. NMF6 strain producing secondary metabolite(s) possessing antimicrobial, antioxidant, anticancer, and antiviral activities. J Genet Eng Biotechnol 19: 102. https://doi.org/10.1186/s43141-021-00203-5
|
| [40] | Kumar V, Bisht GS, Gusain O (2013) Terrestrial actinomycetes from diverse locations of Uttarakhnad, India: Isolation and screening for their antibacterial activity. Iran J Microbiol 5: 299-308. |
| [41] |
Subramani R, Sipkema D (2019) Marine rare Actinomycetes: A Promising source of structurally diverse and unique novel natural products. Mar Drugs 17. https://doi.org/10.3390/md17050249
|
| [42] |
Dhakal D, Pokhrel AR, Shrestha B, et al. (2017) Marine rare Actinobacteria: Isolation, characterization, and strategies for harnessing bioactive compounds. Front Microbiol 8: 1106. https://doi.org/10.3389/fmicb.2017.01106
|
| [43] |
Subramani R, Aalbersberg W (2013) Culturable rare Actinomycetes: diversity, isolation and marine natural product discovery. Appl Microbiol Biotechnol 97: 9291-9321. https://doi.org/10.1007/s00253-013-5229-7
|
| [44] |
Mehbub MF, Lei J, Franco C, et al. (2014) Marine sponge derived natural products between 2001 and 2010: trends and opportunities for discovery of bioactives. Mar Drugs 12: 4539-4577. https://doi.org/10.3390/md12084539
|
| [45] | Sun W, Zhang F, He L, et al. (2015) Actinomycetes from the South China Sea sponges: isolation, diversity, and potential for aromatic polyketides discovery. Front Microbiol 6: 1048. https://doi.org/10.3389/fmicb.2015.01048 |
| [46] |
Tiwari K, Gupta RK (2013) Diversity and isolation of rare actinomycetes: an overview. Crit Rev Microbiol 39: 256-294. https://doi.org/10.3109/1040841X.2012.709819
|
| [47] |
Olson JB, Harmody DK, Bej AK, et al. (2007) Tsukamurella spongiae sp. nov., a novel actinomycete isolated from a deep-water marine sponge. Int J Syst Evol Microbiol 57: 1478-1481. https://doi.org/10.1099/ijs.0.64837-0
|
| [48] |
Siro G, Pipite A, Christi K, et al. (2022) Marine Actinomycetes associated with stony corals: a potential hotspot for specialized metabolites. Microorganisms 10: 1349. https://doi.org/10.3390/microorganisms10071349
|
| [49] |
Girão M, Rego A, Fonseca AC, et al. (2024) Actinomycetota from macroalgae as rich source for natural products discovery revealed through Culture-Dependent and -Independent approaches. Microb Biotechnol 17. https://doi.org/10.1111/1751-7915.70058
|
| [50] |
Schorn MA, Alanjary MM, Aguinaldo K, et al. (2016) Sequencing rare marine actinomycete genomes reveals high density of unique natural product biosynthetic gene clusters. Microbiology (Reading) 162: 2075-2086. https://doi.org/10.1099/mic.0.000386
|
| [51] |
Xi L, Ruan J, Huang Y (2012) Diversity and biosynthetic potential of culturable actinomycetes associated with marine sponges in the China Seas. Int J Mol Sci 13: 5917-5932. https://doi.org/10.3390/ijms13055917
|
| [52] |
Al-Awadhi H, Dashti N, Khanafer M, et al. (2013) Bias problems in culture-independent analysis of environmental bacterial communities: a representative study on hydrocarbonoclastic bacteria. SpringerPlus 2: 369. https://doi.org/10.1186/2193-1801-2-369
|
| [53] |
Vester JK, Glaring MA, Stougaard P (2015) Improved cultivation and metagenomics as new tools for bioprospecting in cold environments. Extremophiles 19: 17-29. https://doi.org/10.1007/s00792-014-0704-3
|
| [54] |
Su C, Lei L, Duan Y, et al. (2012) Culture-independent methods for studying environmental microorganisms: methods, application, and perspective. Appl Microbiol Biotechnol 93: 993-1003. https://doi.org/10.1007/s00253-011-3800-7
|
| [55] |
Cocolin L, Alessandria V, Dolci P, et al. (2013) Culture independent methods to assess the diversity and dynamics of microbiota during food fermentation. Int J Food Microbiol 167: 29-43. https://doi.org/10.1016/j.ijfoodmicro.2013.05.008
|
| [56] |
Maheshwari P, Sankar PM (2023) Chapter 42-Culture-independent and culture-dependent approaches in symbiont analysis: in proteobacteria. Microbial Symbionts . Academic Press 743-763. https://doi.org/10.1016/B978-0-323-99334-0.00018-9
|
| [57] | McCombie WR, McPherson JD, Mardis ER (2019) Next-Generation sequencing technologies. Cold Spring Harb Perspect Med 9. https://doi.org/10.1101/cshperspect.a036798 |
| [58] |
Satam H, Joshi K, Mangrolia U, et al. (2023) Next-Generation sequencing technology: Current trends and advancements. Biology 12: 997. https://doi.org/10.3390/biology12070997
|
| [59] |
Rhoads A, Au KF (2015) PacBio sequencing and its applications. Genomics Proteomics Bioinf 13: 278-289. https://doi.org/10.1016/j.gpb.2015.08.002
|
| [60] |
Bolyen E, Rideout JR, Dillon MR, et al. (2019) Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol 37: 852-857. https://doi.org/10.1038/s41587-019-0209-9
|
| [61] |
Callahan BJ, McMurdie PJ, Rosen MJ, et al. (2016) DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 13: 581-583. https://doi.org/10.1038/nmeth.3869
|
| [62] |
Janssen S, McDonald D, Gonzalez A, et al. (2018) Phylogenetic placement of exact amplicon sequences improves associations with clinical information. mSystems 3: 10.1128/msystems.00021-00018. https://doi.org/10.1128/mSystems.00021-18
|
| [63] |
Faith DP (1992) Conservation evaluation and phylogenetic diversity. Biol Conserv 61: 1-10. https://doi.org/10.1016/0006-3207(92)91201-3
|
| [64] |
Shannon CE (1948) A mathematical theory of communication. Bell Syst Tech J 27: 379-423. https://doi.org/10.1002/j.1538-7305.1948.tb01338.x
|
| [65] |
Lozupone C, Knight R (2005) UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol 71: 8228-8235. https://doi.org/10.1128/AEM.71.12.8228-8235.2005
|
| [66] |
Lozupone CA, Hamady M, Kelley ST, et al. (2007) Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl Environ Microbiol 73: 1576-1585. https://doi.org/10.1128/AEM.01996-06
|
| [67] | Bokulich Nicholas A, Dillon Matthew R, Zhang Y, et al. (2018) q2-longitudinal: Longitudinal and paired-sample analyses of microbiome data. mSystems 3: 10.1128/msystems.00219-00218. https://doi.org/10.1128/msystems.00219-18 |
| [68] | Oren A, Garrity GM (2021) Valid publication of the names of forty-two phyla of prokaryotes. Int J Syst Evol Microbiol 71. https://doi.org/10.1099/ijsem.0.005056 |
| [69] |
Marcelino VR, Clausen P, Buchmann JP, et al. (2020) CCMetagen: comprehensive and accurate identification of eukaryotes and prokaryotes in metagenomic data. Genome Biol 21: 103. https://doi.org/10.1186/s13059-020-02014-2
|
| [70] | Davari N, Jouri M, Ariapour A, et al. (2011) Comparison of measurement indices of diversity, richness, dominance, and evenness in rangeland ecosystem (Case Study: Jvaherdeh-Ramesar). J Rangeland Sci 2: 389-398. |
| [71] |
Zhang H, Zhang W, Jin Y, et al. (2008) A comparative study on the phylogenetic diversity of culturable actinobacteria isolated from five marine sponge species. Antonie van Leeuwenhoek 93: 241-248. https://doi.org/10.1007/s10482-007-9196-9
|
| [72] |
Weeraphan T, Somphong A, Poengsungnoen V, et al. (2023) Bacterial microbiome in tropical lichens and the effect of the isolation method on culturable lichen-derived actinobacteria. Sci Rep 13: 5483. https://doi.org/10.1038/s41598-023-32759-2
|
| [73] | Hall TA (1999) BIOEDIT: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/nt. Nucleic Acids Symposium Series 41: 95-98. |
| [74] |
Chalita M, Kim YO, Park S, et al. (2024) EzBioCloud: a genome-driven database and platform for microbiome identification and discovery. Int J Syst Evol Microbiol 74. https://doi.org/10.1099/ijsem.0.006421
|
| [75] |
McArdle AJ, Kaforou M (2020) Sensitivity of shotgun metagenomics to host DNA: abundance estimates depend on bioinformatic tools and contamination is the main issue. Access Microbiol 2: acmi000104. https://doi.org/10.1099/acmi.0.000104
|
| [76] |
Odendaal ML, Groot JA, Hasrat R, et al. (2022) Higher off-target amplicon detection rate in MiSeq v3 compared to v2 reagent kits in the context of 16S-rRNA-sequencing. Sci Rep 12: 16489. https://doi.org/10.1038/s41598-022-20573-1
|
| [77] | Mohseni M, Norouzi H, Hamedi J, et al. (2013) Screening of antibacterial producing actinomycetes from sediments of the caspian sea. Int J Mol Cell Med 2: 64-71. |
| [78] |
Zhao H, Kassama Y, Young M, et al. (2004) Differentiation of Micromonospora isolates from a coastal sediment in Wales on the basis of Fourier transform infrared spectroscopy, 16S rRNA sequence analysis, and the amplified fragment length polymorphism technique. Appl Environ Microbiol 70: 6619-6627. https://doi.org/10.1128/AEM.70.11.6619-6627.2004
|
| [79] | Putchakarn S, Munhapon A, Taleb S, et al. (2015) Marine sponges in the industrial and conservation areas of Chon Buri and Rayong provinces. Burapha University International Conference 2015 . |
| [80] |
Andreeva NA, Melnikov VV, Snarskaya DD (2020) The role of Cyanobacteria in marine ecosystems. Russ J Marine Biol 46: 154-165. https://doi.org/10.1134/S1063074020030025
|
| [81] |
Pita L, Rix L, Slaby BM, et al. (2018) The sponge holobiont in a changing ocean: from microbes to ecosystems. Microbiome 6: 46. https://doi.org/10.1186/s40168-018-0428-1
|
| [82] |
Kuo J, Yang YT, Lu MC, et al. (2019) Antimicrobial activity and diversity of bacteria associated with Taiwanese marine sponge Theonella swinhoei. Annals Microbiol 69: 253-265. https://doi.org/10.1007/s13213-018-1414-3
|
| [83] |
Mawang CI, Azman AS, Fuad AM, et al. (2021) Actinobacteria: An eco-friendly and promising technology for the bioaugmentation of contaminants. Biotechnol Rep (Amst) 32: e00679. https://doi.org/10.1016/j.btre.2021.e00679
|
| [84] |
Nguyen VH, Wemheuer B, Song W, et al. (2023) Identification, classification, and functional characterization of novel sponge-associated acidimicrobiial species. Syst Appl Microbiol 46: 126426. https://doi.org/10.1016/j.syapm.2023.126426
|
| [85] |
Holert J, Cardenas E, Bergstrand Lee H, et al. (2018) Metagenomes reveal global distribution of bacterial steroid catabolism in natural, engineered, and host environments. mBio 9: 10.1128/mbio.02345-02317. https://doi.org/10.1128/mBio.02345-17
|
| [86] |
Lee YJ, Wagner ID, Brice ME, et al. (2005) Thermosediminibacter oceani gen. nov., sp. nov. and Thermosediminibacter litoriperuensis sp. nov., new anaerobic thermophilic bacteria isolated from Peru Margin. Extremophiles 9: 375-383. https://doi.org/10.1007/s00792-005-0453-4
|
| [87] |
Simister R, Taylor MW, Tsai P, et al. (2012) Thermal stress responses in the bacterial biosphere of the Great Barrier Reef sponge, Rhopaloeides odorabile. Environ Microbiol 14: 3232-3246. https://doi.org/10.1111/1462-2920.12010
|
| [88] |
Hardoim CCP, Lôbo-Hajdu G, Custódio MR, et al. (2021) Prokaryotic, fungal, and unicellular eukaryotic core communities across three sympatric marine sponges from the Southwestern Atlantic Coast are dominated largely by deterministic assemblage processes. Front Microbiol 12: 674004. https://doi.org/10.3389/fmicb.2021.674004
|
| [89] |
Olatunji OO, Petchoubon A, Thawai C, et al. (2020) Allocation of trisoxazole macrolides in the sponge Penares cf. nux and the impact on epiphytic bacterial cohabitants. Nat Prod Res 34: 710-713. https://doi.org/10.1080/14786419.2018.1493589
|
| [90] |
Santalova EA, Kolesnikova SA (2024) 9-Chloro-5,9-dienoic and other fatty acids from marine sponge Penares sp. Russ J Bioorg Chem 50: 418-431. https://doi.org/10.1134/S1068162024020249
|
| [91] | Back CR, Stennett HL, Williams SE, et al. (2021) A new micromonospora strain with antibiotic activity isolated from the microbiome of a Mid-Atlantic Deep-Sea Sponge. Mar Drugs 19. https://doi.org/10.3390/md19020105 |
| [92] | Esteves AI, Cullen A, Thomas T (2017) Competitive interactions between sponge-associated bacteria. FEMS Microbiol Ecol 93. https://doi.org/10.1093/femsec/fix008 |
| [93] |
Songsumanus A, Tanasupawat S, Igarashi Y, et al. (2013) Micromonospora maritima sp. nov., isolated from mangrove soil. Int J Syst Evol Microbiol 63: 554-559. https://doi.org/10.1099/ijs.0.039180-0
|
| [94] |
Yan S, Zeng M, Wang H, et al. (2022) Micromonospora: A Prolific source of bioactive secondary metabolites with therapeutic potential. J Med Chem 65: 8735-8771. https://doi.org/10.1021/acs.jmedchem.2c00626
|
| [95] | Waleed MA, Mousa AA (2016) Enzymes and phytohormones from Micromonospora. Actinobacteria . Rijeka: IntechOpen. Ch. 12 |
| [96] |
Li L, Zhu H, Xu Q, et al. (2019) Micromonospora craniellae sp. nov., isolated from a marine sponge, and reclassification of Jishengella endophytica as Micromonospora endophytica comb. nov. Int J Syst Evol Microbiol 69: 715-720. https://doi.org/10.1099/ijsem.0.003209
|
| [97] | Jose PA, Jha B (2017) Intertidal marine sediment harbours Actinobacteria with promising bioactive and biosynthetic potential. Sci Rep 7. https://doi.org/10.1038/s41598-017-09672-6 |
| [98] |
Hassane CS, Fouillaud M, Goff GL, et al. (2020) Microorganisms associated with the marine sponge Scopalina hapalia: A reservoir of bioactive molecules to slow down the aging process. Microorganisms 8: 1262. https://doi.org/10.3390/microorganisms8091262
|
| [99] |
Jensen PR, Mafnas C (2006) Biogeography of the marine actinomycete. Salinispora Environ Microbiol 8: 1881-1888. https://doi.org/10.1111/j.1462-2920.2006.01093.x
|
| [100] |
Miksch S, Meiners M, Meyerdierks A, et al. (2021) Bacterial communities in temperate and polar coastal sands are seasonally stable. ISME Commun 1. https://doi.org/10.1038/s43705-021-00028-w
|
 microbiol-11-01-010-s001.pdf microbiol-11-01-010-s001.pdf |
 |
| microbiol-11-01-010-s002.xlsx |
|
Figures(5) / Tables(1)

Trinset Weeraphan, Chollabuppha Chou, Naphatson Chanthathamrongsiri, Thanchanok Sirirak, Sumaitt Putchakarn, Supakarn Chamni, Wongsakorn Phongsopitanun. Microbiome profiling and Actinomycetes isolation from tropical marine sponges[J]. AIMS Microbiology, 2025, 11(1): 182-205. doi: 10.3934/microbiol.2025010







 DownLoad:
DownLoad: