Several pathological conditions might cause the degradation of the cyclin-dependent kinase inhibitor (CKI) p27 and cell cycle arrest at the G1 phase, including cancers and infections. Chlamydia trachomatis (Ctr), as an obligatory intracellular pathogen, has been found to alter the fate of the cell from different aspects. In this study, we aimed to investigate the effect of Ctr infection on the expression of the important cell cycle regularity protein p27 in mesenchymal stem cells (MSCs).
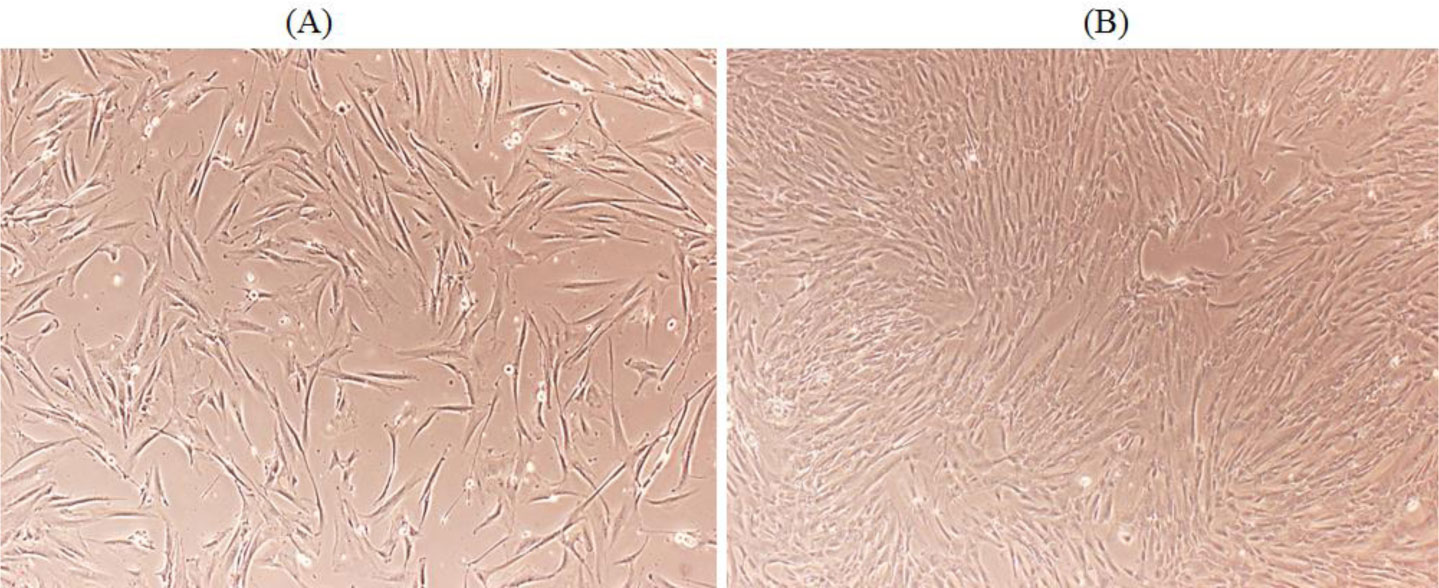
Isolation of MSCs from healthy human fallopian tube was confirmed by detection of the stemness markers Sox2, Nanog and Oct4 and the surface markers CD44, CD73 and CD90 by Western blotting and fluorescence-activated cell sorting analysis. The expression of p27 was downregulated at the protein level upon Ctr D infection measured by Real-Time Quantitative Reverse Transcription PCR (qRT-PCR), IF and Western blotting. Recovery of p27 in Ctr D-infected MSCs was achieved by treatment with difluoromethylornithine (DFMO). Ctr D infected MSCs were able to produce colonies in anchorage-independent soft agar assay.
Ctr D infection was able to downregulate the expression of the important cell cycle regulator protein p27, which will be considered a putative candidate for transformation in Ctr D infected MSCs.
Citation: Mohammad A. Abu-Lubad, Wael Al-Zereini, Munir A. Al-Zeer. Deregulation of the cyclin-dependent kinase inhibitor p27 as a putative candidate for transformation in Chlamydia trachomatis infected mesenchymal stem cells[J]. AIMS Microbiology, 2023, 9(1): 131-150. doi: 10.3934/microbiol.2023009
Several pathological conditions might cause the degradation of the cyclin-dependent kinase inhibitor (CKI) p27 and cell cycle arrest at the G1 phase, including cancers and infections. Chlamydia trachomatis (Ctr), as an obligatory intracellular pathogen, has been found to alter the fate of the cell from different aspects. In this study, we aimed to investigate the effect of Ctr infection on the expression of the important cell cycle regularity protein p27 in mesenchymal stem cells (MSCs).
Isolation of MSCs from healthy human fallopian tube was confirmed by detection of the stemness markers Sox2, Nanog and Oct4 and the surface markers CD44, CD73 and CD90 by Western blotting and fluorescence-activated cell sorting analysis. The expression of p27 was downregulated at the protein level upon Ctr D infection measured by Real-Time Quantitative Reverse Transcription PCR (qRT-PCR), IF and Western blotting. Recovery of p27 in Ctr D-infected MSCs was achieved by treatment with difluoromethylornithine (DFMO). Ctr D infected MSCs were able to produce colonies in anchorage-independent soft agar assay.
Ctr D infection was able to downregulate the expression of the important cell cycle regulator protein p27, which will be considered a putative candidate for transformation in Ctr D infected MSCs.

| [1] |
Huai P, Li F, Chu T, et al. (2020) Prevalence of genital Chlamydia trachomatis infection in the general population: a meta-analysis. BMC Infect Dis 20: 1-8. https://doi.org/10.1186/s12879-020-05307-w 
|
| [2] |
Herweg JA, Rudel T (2016) Interaction of Chlamydiae with human macrophages. The FEBS J 283: 608-618. https://doi.org/10.1111/febs.13609
|
| [3] |
Scherler A, Jacquier N, Kebbi-Beghdadi C, et al. (2020) Diverse stress-inducing treatments cause distinct aberrant body morphologies in the Chlamydia-related bacterium, Waddlia chondrophila. Microorganisms 8: 89. https://doi.org/10.3390/microorganisms8010089
|
| [4] |
Miyairi I, Mahdi OS, Ouellette SP, et al. (2006) Different growth rates of Chlamydia trachomatis biovars reflect pathotype. J Infect Dis 194: 350-357. https://doi.org/10.1086/505432
|
| [5] |
Wilson D, Mathews S, Wan C, et al. (2004) Use of a quantitative gene expression assay based on micro-array techniques and a mathematical model for the investigation of chlamydial generation time. Bull Math Biology 66: 523-537. https://doi.org/10.1016/j.bulm.2003.09.001
|
| [6] | Malhotra M, Sood S, Mukherjee A, et al. (2013) Genital Chlamydia trachomatis: an update. Indian J Med Res 138: 303-316. |
| [7] | Zielecki J Establishment of in vitro-infection models for Chlamydia trachomatis based on human primary cells and primary tissue (2011). |
| [8] |
Abu-Lubad M, Meyer TF, Al-Zeer MA (2014) Chlamydia trachomatis inhibits inducible NO synthase in human mesenchymal stem cells by stimulating polyamine synthesis. J Immunol 193: 2941-2951. https://doi.org/10.4049/jimmunol.1400377
|
| [9] | Orciani M, Caffarini M, Lazzarini R, et al. (2018) Mesenchymal stem cells from cervix and age: new insights into CIN regression rate. Oxi Med Cell Longevity 2018: 1545784. https://doi.org/10.1155/2018/1545784 |
| [10] |
Yan Z, Guo F, Yuan Q, et al. (2019) Endometrial mesenchymal stem cells isolated from menstrual blood repaired epirubicin-induced damage to human ovarian granulosa cells by inhibiting the expression of Gadd45b in cell cycle pathway. Stem Cell Res Ther 10: 1-10. https://doi.org/10.1186/s13287-018-1101-0
|
| [11] |
Bishop RC, Boretto M, Rutkowski MR, et al. (2020) Murine endometrial organoids to model Chlamydia infection. Front Cell Infect Microbiol 10: 416. https://doi.org/10.3389/fcimb.2020.00416
|
| [12] |
Chan RW, Schwab KE, Gargett CE (2004) Clonogenicity of human endometrial epithelial and stromal cells. Biolo Reprod 70: 1738-1750. https://doi.org/10.1095/biolreprod.103.024109
|
| [13] |
Lu J, Zhu L, Zhang L, et al. (2017) Abnormal expression of TRAIL receptors in decidual tissue of Chlamydia trachomatis-infected rats during early pregnancy loss. Reprod Sci 24: 1041-1052. https://doi.org/10.1177/1933719116676393
|
| [14] |
Gargett CE (2007) Uterine stem cells: what is the evidence?. Hum Reprod Update 13: 87-101. https://doi.org/10.1093/humupd/dml045
|
| [15] |
Jazedje T, Perin PM, Czeresnia CE, et al. (2009) Human fallopian tube: a new source of multipotent adult mesenchymal stem cells discarded in surgical procedures. J Transl Med 7: 1-10. https://doi.org/10.1186/1479-5876-7-46
|
| [16] |
Xia M, Bumgarner RE, Lampe MF, et al. (2003) Chlamydia trachomatis infection alters host cell transcription in diverse cellular pathways. J Infect Dis 187: 424-434. https://doi.org/10.1086/367962
|
| [17] |
Al-Zeer MA, Al-Younes HM, Lauster D, et al. (2013) Autophagy restricts Chlamydia trachomatis growth in human macrophages via IFNG-inducible guanylate binding proteins. Autophagy 9: 50-62. https://doi.org/10.4161/auto.22482
|
| [18] |
Al-Zeer MA, Xavier A, Lubad MA, et al. (2017) Chlamydia trachomatis prevents apoptosis via activation of PDPK1-MYC and enhanced mitochondrial binding of hexokinase II. EBioMedicine 23: 100-110. https://doi.org/10.1016/j.ebiom.2017.08.005
|
| [19] |
Balsara ZR, Misaghi S, Lafave JN, et al. (2006) Chlamydia trachomatis infection induces cleavage of the mitotic cyclin B1. Infect Immun 74: 5602-5608. https://doi.org/10.1128/IAI.00266-06
|
| [20] |
Fischer SF, Vier J, Kirschnek S, et al. (2004) Chlamydia inhibit host cell apoptosis by degradation of proapoptotic BH3-only proteins. J Exp Med 200: 905-916. https://doi.org/10.1084/jem.20040402
|
| [21] | González E, Rother M, Kerr MC, et al. (2014) Chlamydia infection depends on a functional MDM2-p53 axis. Nat Commun 5: 1-10. https://doi.org/10.1038/ncomms6201 |
| [22] |
García-Gutiérrez L, Bretones G, Molina E, et al. (2019) Myc stimulates cell cycle progression through the activation of Cdk1 and phosphorylation of p27. Sci Rep 9: 1-17. https://doi.org/10.1038/s41598-019-54917-1
|
| [23] |
Kawauchi T (2014) C dk5 regulates multiple cellular events in neural development, function and disease. Dev Growth Differ 56: 335-348. https://doi.org/10.1111/dgd.12138
|
| [24] |
Niu Y, Xu J, Sun T (2019) Cyclin-dependent kinases 4/6 inhibitors in breast cancer: current status, resistance, and combination strategies. J Cancer 10: 5504. https://doi.org/10.7150/jca.32628
|
| [25] |
Asghar U, Witkiewicz AK, Turner NC, et al. (2015) The history and future of targeting cyclin-dependent kinases in cancer therapy. Nat Rev Drug Discovery 14: 130-146. https://doi.org/10.1038/nrd4504
|
| [26] |
Larrea MD, Wander SA, Slingerland J (2009) p27 as Jekyll and Hyde: regulation of cell cycle and cell motility. Cell Cycle 8: 3455-3461. https://doi.org/10.4161/cc.8.21.9789
|
| [27] |
Alkarain A, Slingerland J (2003) Deregulation of p27 by oncogenic signaling and its prognostic significance in breast cancer. Breast Cancer Res 6: 1-9. https://doi.org/10.1186/bcr722
|
| [28] |
Zhao D, Besser AH, Wander SA, et al. (2015) Cytoplasmic p27 promotes epithelial–mesenchymal transition and tumor metastasis via STAT3-mediated Twist1 upregulation. Oncogene 34: 5447-5459. https://doi.org/10.1038/onc.2014.473
|
| [29] |
Eguchi H, Carpentier S, Kim S, et al. (2004) P27kip1 regulates the apoptotic response of gastric epithelial cells to Helicobacter pylori. Gut 53: 797-804. https://doi.org/10.1136/gut.2003.032144
|
| [30] |
Kim SS, Meitner P, Konkin TA, et al. (2006) Altered expression of Skp2, c-Myc and p27 proteins but not mRNA after H. pylori eradication in chronic gastritis. Mod Pathol 19: 49-58. https://doi.org/10.1038/modpathol.3800476
|
| [31] |
Yarmishyn A, Child ES, Elphick LM, et al. (2008) Differential regulation of the cyclin-dependent kinase inhibitors p21Cip1 and p27Kip1 by phosphorylation directed by the cyclin encoded by Murine Herpesvirus 68. Exp Cell Res 314: 204-212. https://doi.org/10.1016/j.yexcr.2007.09.016
|
| [32] |
Munder M (2009) Arginase: an emerging key player in the mammalian immune system. Br J Pharmacol 158: 638-651. https://doi.org/10.1111/j.1476-5381.2009.00291.x
|
| [33] |
Choi SH, Kim SW, Choi DH, et al. (2000) Polyamine-depletion induces p27Kip1 and enhances dexamethasone-induced G1 arrest and apoptosis in human T lymphoblastic leukemia cells. Leuk Res 24: 119-127. https://doi.org/10.1016/S0145-2126(99)00161-7
|
| [34] |
Tabib A, Bachrach U (1998) Polyamines induce malignant transformation in cultured NIH 3T3 fibroblasts. Int J Biochem Cell Biol 30: 135-146. https://doi.org/10.1016/S1357-2725(97)00073-3
|
| [35] | Ravanko K, Järvinen K, Paasinen-Sohns A, et al. (2000) Loss of p27Kip1 from cyclin E/cyclin-dependent kinase (CDK) 2 but not from cyclin D1/CDK4 complexes in cells transformed by polyamine biosynthetic enzymes. Cancer Res 60: 5244-5253. |
| [36] |
Koomoa D-LT, Geerts D, Lange I, et al. (2013) DFMO/eflornithine inhibits migration and invasion downstream of MYCN and involves p27Kip1 activity in neuroblastoma. Int J Oncol 42: 1219-1228. https://doi.org/10.3892/ijo.2013.1835
|
| [37] |
Al-Younes HM, Rudel T, Brinkmann V, et al. (2001) Low iron availability modulates the course of Chlamydia pneumoniae infection. Cell Microbiol 3: 427-437. https://doi.org/10.1046/j.1462-5822.2001.00125.x
|
| [38] |
Hristova NR, Tagscherer KE, Fassl A, et al. (2013) Notch1-dependent regulation of p27 determines cell fate in colorectal cancer. Intl J Oncol 43: 1967-1975. https://doi.org/10.3892/ijo.2013.2140
|
| [39] |
Zhu H, Shen Z, Luo H, et al. (2016) Chlamydia trachomatis infection-associated risk of cervical cancer: a meta-analysis. Medicine 95: e3077. https://doi.org/10.1097/MD.0000000000003077
|
| [40] |
Stone KM, Zaidi A, Rosero-Bixby L, et al. (1995) Sexual behavior, sexually transmitted diseases, and risk of cervical cancer. Epidemiology : 409-414. https://doi.org/10.1097/00001648-199507000-00014
|
| [41] |
Farivar TN, Johari P (2012) Lack of association between Chlamydia trachomatis infection and cervical cancer-Taq Man realtime PCR assay findings. Asian Pac J Cancer Prev 13: 3701-3704. https://doi.org/10.7314/APJCP.2012.13.8.3701
|
| [42] |
Tungsrithong N, Kasinpila C, Maneenin C, et al. (2014) Lack of significant effects of Chlamydia trachomatis infection on cervical cancer risk in a nested case-control study in North-East Thailand. Asian Pac J Cancer Prev 15: 1497-1500. https://doi.org/10.7314/APJCP.2014.15.3.1497
|
| [43] |
Zereu M, Zettler C, Cambruzzi E, et al. (2007) Herpes simplex virus type 2 and Chlamydia trachomatis in adenocarcinoma of the uterine cervix. Gynecol Oncol 105: 172-175. https://doi.org/10.1016/j.ygyno.2006.11.006
|
| [44] |
Simonetti AC, de Lima Melo JH, de Souza PRE, et al. (2009) Immunological's host profile for HPV and Chlamydia trachomatis, a cervical cancer cofactor. Microbes Infect 11: 435-442. https://doi.org/10.1016/j.micinf.2009.01.004
|
| [45] |
Gunin AG, Glyakin DS, Emelianov VU (2021) Mycoplasma and Chlamydia infection can increase risk of endometrial cancer by pro-inflammatory cytokine enlargement. Indian J Gynecol Oncol 19: 1-7. https://doi.org/10.1007/s40944-020-00477-6
|
| [46] | Parazzini F, La Vecchia C, Negri E, et al. (1996) Pelvic inflammatory disease and risk of ovarian cancer. Cancer Epidemiol Biomarkers Prve 5: 667-669. |
| [47] |
Richards TS, Knowlton AE, Grieshaber SS (2013) Chlamydia trachomatis homotypic inclusion fusion is promoted by host microtubule trafficking. BMC Microbiol 13: 1-8. https://doi.org/10.1186/1471-2180-13-185
|
| [48] |
Koskela P, Anttila T, Bjørge T, et al. (2000) Chlamydia trachomatis infection as a risk factor for invasive cervical cancer. Int J Cancer 85: 35-39. https://doi.org/10.1002/(SICI)1097-0215(20000101)85:1<35::AID-IJC6>3.0.CO;2-A
|
| [49] |
Smith JS, Bosetti C, MUnoz N, et al. (2004) Chlamydia trachomatis and invasive cervical cancer: A pooled analysis of the IARC multicentric case-control study. Int J Cancer 111: 431-439. https://doi.org/10.1002/ijc.20257
|
| [50] |
Smith JS, Muñoz N, Herrero R, et al. (2002) Evidence for Chlamydia trachomatis as a human papillomavirus cofactor in the etiology of invasive cervical cancer in Brazil and the Philippines. J Infect Dis 185: 324-331. https://doi.org/10.1086/338569
|
| [51] |
Hinkula M, Pukkala E, Kyyrönen P, et al. (2004) A population-based study on the risk of cervical cancer and cervical intraepithelial neoplasia among grand multiparous women in Finland. Bri J Cancer 90: 1025-1029. https://doi.org/10.1038/sj.bjc.6601650
|
| [52] |
Lindahl T, Barnes D (2000) Repair of endogenous DNA damage. Cold Spring Harb Symp Quant Biol 65: 127-134. https://doi.org/10.1101/sqb.2000.65.127
|
| [53] |
Coombes BK, Mahony JB (1999) Chlamydia pneumoniae infection of human endothelial cells induces proliferation of smooth muscle cells via an endothelial cell-derived soluble factor (s). Infect Immun 67: 2909-2915. https://doi.org/10.1128/IAI.67.6.2909-2915.1999
|
| [54] |
Chiou C-C, Chan C-C, Kuo Y-P, et al. (2003) Helicobacter pylori inhibits activity of cdc2 kinase and delays G 2/M to G 1 progression in gastric adenocarcinoma cell. Scand J Gastroenterol 38: 147-152. https://doi.org/10.1080/00365520310000627
|
| [55] |
Nougayrède J-P, Boury M, Tasca C, et al. (2001) Type III secretion-dependent cell cycle block caused in HeLa cells by enteropathogenic Escherichia coli O103. Infect Immun 69: 6785-6795. https://doi.org/10.1128/IAI.69.11.6785-6795.2001
|
| [56] |
Davy CE, Jackson DJ, Raj K, et al. (2005) Human papillomavirus type 16 E1∧ E4-induced G2 arrest is associated with cytoplasmic retention of active Cdk1/cyclin B1 complexes. J Virol 79: 3998-4011. https://doi.org/10.1128/JVI.79.7.3998-4011.2005
|
| [57] |
Poggioli GJ, Dermody TS, Tyler KL (2001) Reovirus-induced ς1s-dependent G2/M phase cell cycle arrest is associated with inhibition of p34cdc2. J Virol 75: 7429-7434. https://doi.org/10.1128/JVI.75.16.7429-7434.2001
|
| [58] |
He J, Choe S, Walker R, et al. (1995) Human immunodeficiency virus type 1 viral protein R (Vpr) arrests cells in the G2 phase of the cell cycle by inhibiting p34cdc2 activity. J Virol 69: 6705-6711. https://doi.org/10.1128/jvi.69.11.6705-6711.1995
|
| [59] |
Slingerland J, Pagano M (2000) Regulation of the cdk inhibitor p27 and its deregulation in cancer. J Cell Physiol 183: 10-17. https://doi.org/10.1002/(SICI)1097-4652(200004)183:1<10::AID-JCP2>3.0.CO;2-I
|
| [60] |
Porter PL, Malone KE, Heagerty PJ, et al. (1997) Expression of cell-cycle regulators p27Kip1 and cyclin E, alone and in combination, correlate with survival in young breast cancer patients. Nat Med 3: 222-225. https://doi.org/10.1038/nm0297-222
|
| [61] |
Catzavelos C, Bhattacharya N, Ung YC, et al. (1997) Decreased levels of the cell-cycle inhibitor p27Kip1 protein: prognostic implications in primary breast cancer. Nat Med 3: 227-230. https://doi.org/10.1038/nm0297-227
|
| [62] |
Nakayama K, Ishida N, Shirane M, et al. (1996) Mice lacking p27Kip1 display increased body size, multiple organ hyperplasia, retinal dysplasia, and pituitary tumors. Cell 85: 707-720. https://doi.org/10.1016/S0092-8674(00)81237-4
|
| [63] |
Fujita N, Sato S, Katayama K, et al. (2002) Akt-dependent phosphorylation of p27Kip1 promotes binding to 14-3-3 and cytoplasmic localization. J Biol Chem 277: 28706-28713. https://doi.org/10.1074/jbc.M203668200
|
| [64] |
Tsvetkov LM, Yeh K-H, Lee S-J, et al. (1999) p27Kip1 ubiquitination and degradation is regulated by the SCFSkp2 complex through phosphorylated Thr187 in p27. Curr Biol 9: 661-S662. https://doi.org/10.1016/S0960-9822(99)80290-5
|
| [65] |
Hengst L, Reed SI (1996) Translational control of p27Kip1 accumulation during the cell cycle. Science 271: 1861-1864. https://doi.org/10.1126/science.271.5257.1861
|
| [66] | Eguchi H, Herschenhous N, Kuzushita N, et al. (2003) Helicobacter pylori increases proteasome-mediated degradation of p27kip1 in gastric epithelial cells. Cancer Res 63: 4739-4746. |
| [67] |
Wen S, So Y, Singh K, et al. (2012) Promotion of cytoplasmic mislocalization of p27 by Helicobacter pylori in gastric cancer. Oncogene 31: 1771-1780. https://doi.org/10.1038/onc.2011.362
|
| [68] |
Sekimoto T, Fukumoto M, Yoneda Y (2004) 14-3-3 suppresses the nuclear localization of threonine 157-phosphorylated p27Kip1. EMBO J 23: 1934-1942. https://doi.org/10.1038/sj.emboj.7600198
|
| [69] |
Ishida N, Hara T, Kamura T, et al. (2002) Phosphorylation of p27Kip1 on serine 10 is required for its binding to CRM1 and nuclear export. J Biol Chem 277: 14355-14358. https://doi.org/10.1074/jbc.C100762200
|
| [70] |
Kotoshiba S, Gopinathan L, Pfeiffenberger E, et al. (2014) p27 is regulated independently of Skp2 in the absence of Cdk2. Biochim Biophy Acta Mol Cell Res 1843: 436-445. https://doi.org/10.1016/j.bbamcr.2013.11.005
|
| [71] |
Short JD, Dere R, Houston KD, et al. (2010) AMPK-mediated phosphorylation of murine p27 at T197 promotes binding of 14-3-3 proteins and increases p27 stability. Mol Carcinog 49: 429-439. https://doi.org/10.1002/mc.20613
|
| [72] |
Koomoa D-LT, Yco LP, Borsics T, et al. (2008) Ornithine decarboxylase inhibition by α-difluoromethylornithine activates opposing signaling pathways via phosphorylation of both Akt/protein kinase B and p27Kip1 in neuroblastoma. Cancer Res 68: 9825-9831. https://doi.org/10.1158/0008-5472.CAN-08-1865
|
| [73] |
Martín A, Odajima J, Hunt SL, et al. (2005) Cdk2 is dispensable for cell cycle inhibition and tumor suppression mediated by p27Kip1 and p21Cip1. Cancer Cell 7: 591-598. https://doi.org/10.1016/j.ccr.2005.05.006
|
| [74] | Krysenko S, Wohlleben W (2022) Polyamine and ethanolamine metabolism in bacteria as an important component of nitrogen assimilation for survival and pathogenicity. Med Sci 10: 40. https://doi.org/10.3390/medsci10030040 |
| [75] |
Knowlton AE, Fowler LJ, Patel RK, et al. (2013) Chlamydia induces anchorage independence in 3T3 cells and detrimental cytological defects in an infection model. PLOS One 8: e54022. https://doi.org/10.1371/journal.pone.0054022
|
| [76] |
Kumar A, Tripathy MK, Pasquereau S, et al. (2018) The human cytomegalovirus strain DB activates oncogenic pathways in mammary epithelial cells. EBioMedicine 30: 167-183. https://doi.org/10.1016/j.ebiom.2018.03.015
|
| [77] |
Chen AL, Johnson KA, Lee JK, et al. (2012) CPAF: a Chlamydial protease in search of an authentic substrate. PLoS Pathog 8: e1002842. https://doi.org/10.1371/journal.ppat.1002842
|








Figures(9)

Mohammad A. Abu-Lubad, Wael Al-Zereini, Munir A. Al-Zeer. Deregulation of the cyclin-dependent kinase inhibitor p27 as a putative candidate for transformation in Chlamydia trachomatis infected mesenchymal stem cells[J]. AIMS Microbiology, 2023, 9(1): 131-150. doi: 10.3934/microbiol.2023009







 DownLoad:
DownLoad: