Citation: Thomas Rohrlack. Low temperatures can promote cyanobacterial bloom formation by providing refuge from microbial antagonists[J]. AIMS Microbiology, 2018, 4(2): 304-318. doi: 10.3934/microbiol.2018.2.304

| [1] |
Paerl HW, Otten TG (2013) Harmful cyanobacterial blooms: causes, consequences, and controls. Microbial Ecol 65: 995–1010. doi: 10.1007/s00248-012-0159-y

|
| [2] | Ibelings BW, Fastner J, Bormans M, et al. (2016) Cyanobacterial blooms. Ecology, prevention, mitigation and control: Editorial to a CYANOCOST Special Issue. Aquat Ecol 50: 327–331. |
| [3] |
Van Wichelen J, Vanormelingen P, Codd GA, et al. (2016) The common bloom-forming cyanobacterium Microcystis is prone to a wide array of microbial antagonists. Harmful Algae 55: 97–111. doi: 10.1016/j.hal.2016.02.009
|
| [4] |
Gerphagnon M, Macarthur DJ, Latour D, et al. (2015) Microbial players involved in the decline of filamentous and colonial cyanobacterial blooms with a focus on fungal parasitism. Environ Microbiol 17: 2573–2587. doi: 10.1111/1462-2920.12860
|
| [5] |
Thingstad TF, Lignell R (1997) Theoretical models for the control of bacterial growth rate, abundance, diversity and carbon demand. Aquat Microb Ecol 13: 19–27. doi: 10.3354/ame013019
|
| [6] |
Kuno S, Yoshida T, Kaneko T, et al. (2012) Intricate Interactions between the bloom-forming cyanobacterium Microcystis aeruginosa and foreign genetic elements, revealed by diversified clustered regularly interspaced short palindromic repeat (CRISPR) signatures. Appl Environ Microbiol 78: 5353–5360. doi: 10.1128/AEM.00626-12
|
| [7] |
Rohrlack T, Christiansen G, Kurmayer R (2013) Putative antiparasite defensive system involving ribosomal and nonribosomal oligopeptides in cyanobacteria of the Genus Planktothrix. Appl Environ Microbiol 79: 2642–2647. doi: 10.1128/AEM.03499-12
|
| [8] |
Pajdak-Stos A, Fialkowska E, Fyda J (2001) Phormidium autumnale (Cyanobacteria) defense against three ciliate grazer species. Aquat Microb Ecol 23: 237–244. doi: 10.3354/ame023237
|
| [9] |
Agha R, Lezcano MA, Labrador MDM, et al. (2014) Seasonal dynamics and sedimentation patterns of Microcystis oligopeptide-based chemotypes reveal subpopulations with different ecological traits. Limnol Oceanogr 59: 861–871. doi: 10.4319/lo.2014.59.3.0861
|
| [10] |
Sønstebø JH, Rohrlack T (2011) Possible implications of chytrid parasitism for population subdivision in freshwater cyanobacteria of the genus Planktothrix. Appl Environ Microbiol 77: 1344–1351. doi: 10.1128/AEM.02153-10
|
| [11] |
Jenkins CA, Hayes PK (2006) Diversity of cyanophages infecting the heterocystous filamentous cyanobacterium Nodularia isolated from the brackish Baltic Sea. J Mar Biol Assoc UK 86: 529–536. doi: 10.1017/S0025315406013439
|
| [12] |
Kimura S, Sako Y, Yoshida T (2013) Rapid Microcystis cyanophage gene diversification revealed by long- and short-term genetic analyses of the tail sheath gene in a natural pond. Appl Environ Microbiol 79: 2789–2795. doi: 10.1128/AEM.03751-12
|
| [13] | Brockhurst MA, Chapman T, King KC, et al. (2014) Running with the Red Queen: the role of biotic conflicts in evolution. P Roy Soc B-Biol Sci 281: 9. |
| [14] |
Scholthof KBG (2007) The disease triangle: pathogens, the environment and society. Nat Rev Microbiol 5: 152–156. doi: 10.1038/nrmicro1596
|
| [15] |
Chiaramonte LV, Ray RA, Corum RA, et al. (2016) Klamath River thermal refuge provides juvenile salmon reduced exposure to the parasite Ceratonova shasta. T Am Fish Soc 145: 810–820. doi: 10.1080/00028487.2016.1159612
|
| [16] | Tobler M, Schlupp I, de Leon FJG, et al. (2007) Extreme habitats as refuge from parasite infections? Evidence from an extremophile fish. Acta Oecol 31: 270–275. |
| [17] |
Strom SL, Harvey EL, Fredrickson KA, et al. (2013) Broad salinity tolerance as a refuge from predation in the harmful raphidophyte alga Heterosigma akashiwo (Raphidophyceae). J Phycol 49: 20–31. doi: 10.1111/jpy.12013
|
| [18] | Bruning K (1991) Infection of the diatom Asterionella by a chytrid. II. Effects of light on survival and epidemic development of the parasite. J Plankton Res 13: 119–129. |
| [19] |
Bruning K (1991) Effect of phosphorus limitation on the epidemiology of a chytrid phytoplankton parasite. Freshwater Biol 25: 409–417. doi: 10.1111/j.1365-2427.1991.tb01385.x
|
| [20] | Gsell AS, Domis LND, van Donk E, et al. (2013) Temperature alters host genotype-specific susceptibility to chytrid infection. PLoS One 8: 10. |
| [21] |
Bruning K (1991) Effects of temperature and light on the population-dynamics of the Asterionella-Rhizophydium association. J Plankton Res 13: 707–719. doi: 10.1093/plankt/13.4.707
|
| [22] |
Ibelings BW, Gsell AS, Mooij WM, et al. (2011) Chytrid infections and diatom spring blooms: paradoxical effects of climate warming on fungal epidemics in lakes. Freshwater Biol 56: 754–766. doi: 10.1111/j.1365-2427.2010.02565.x
|
| [23] |
Mankiewicz-Boczek J, Jaskulska A, Paweczyk J, et al. (2016) Cyanophages infection of Microcystis bloom in lowland dam reservoir of Sulejw, Poland. Microbial Ecol 71: 315–325. doi: 10.1007/s00248-015-0677-5
|
| [24] | Gerphagnon M, Colombet J, Latour D, et al. (2017) Spatial and temporal changes of parasitic chytrids of cyanobacteria. Sci Rep UK 7. |
| [25] |
Gleason FH, Lilje O (2009) Structure and function of fungal zoospores: ecological implications. Fungal Ecol 2: 53–59. doi: 10.1016/j.funeco.2008.12.002
|
| [26] |
Lindstedt KJ (1971) Chemical control of feeding behavior. Comp Biochem Phys A 39: 553–581. doi: 10.1016/0300-9629(71)90319-7
|
| [27] | Berge DM, Løvik JE, Brettum P (1985) Tyrifjordenundersøkelsen 1978–1981, Datarapport. NIVA Rep 1777: 249. |
| [28] | Halstvedt CB, Rohrlack T, Andersen T, et al. (2007) Seasonal dynamics and depth distribution of Planktothrix spp. in Lake Steinsfjorden (Norway) related to environmental factors. J Plankton Res 29: 471–482. |
| [29] | Beard SJ, Davis PA, Iglesias-Rodriguez D, et al. (2000) Gas Vesicle genes in Planktothrix spp. from Nordic lakes: strains with weak gas vesicles possess a longer variant of gvpC. Microbiology 146: 2009–2018. |
| [30] |
Perga ME, Domaizon I, Guillard J, et al. (2013) Are cyanobacterial blooms trophic dead ends? Oecologia 172: 551–562. doi: 10.1007/s00442-012-2519-1
|
| [31] |
Kurmayer R, Juttner F (1999) Strategies for the co-existence of zooplankton with the toxic cyanobacterium Planktothrix rubescens in Lake Zurich. J Plankton Res 21: 659–683. doi: 10.1093/plankt/21.4.659
|
| [32] | Gliwicz ZM (1990) Why do cladocerans fail to control algal blooms? Hydrobiologia 200: 83–97. |
| [33] | Kyle M (2015) Learning from the past; using lake sediments as chemical and biological archives. Norwegian University of Life Sciences. |
| [34] |
Canter HM, Heaney SI, Lund JWG (1990) The ecological significance of grazing on plnaktonic populations od cyanobacteria by the ciliate Nassula. New Phytol 114: 247–263. doi: 10.1111/j.1469-8137.1990.tb00397.x
|
| [35] | Davis PA, Dent M, Parker J, et al. (2003) The annual cycle of growth rate and biomass change in Planktothrix spp. in Blelham Tarn, English Lake District. Freshwater Biol 48: 852–867. |
| [36] |
Brabrand A, Faafeng BA, Kallqvist T, et al. (1983) Biological control of undesirable cyanobacteria in culturally eutrohpic lakes. Oecologia 60: 1–5. doi: 10.1007/BF00379311
|
| [37] |
Hausmann K, Ruskens A (1984) Studies on the digestion in the ciliate Nassula aurea Ehrenberg. Arch Protistenkd 128: 77–87. doi: 10.1016/S0003-9365(84)80030-5
|
| [38] |
Rohrlack T, Edvardsen B, Skulberg R, et al. (2008) Oligopeptide chemotypes of the toxic freshwater cyanobacterium Planktothrix can form subpopulations with dissimilar ecological traits. Limnol Oceanogr 53: 1279–1293. doi: 10.4319/lo.2008.53.4.1279
|
| [39] |
Rounge TB, Rohrlack T, Decenciere B, et al. (2010) Subpopulation differentiation associated with nonribosomal peptide synthese gene cluster dynamics in the cyanobacterium Planktothrix spp. J Phycol 46: 645–652. doi: 10.1111/j.1529-8817.2010.00856.x
|
| [40] |
Rounge TB, Rohrlack T, Nederbragt AJ, et al. (2009) A genome-wide analysis of nonribosomal peptide synthetase gene clusters and their peptides in a Planktothrix rubescens strain. BMC Genomics 10: 396. doi: 10.1186/1471-2164-10-396
|
| [41] |
Tooming-Klunderud A, Sogge H, Rounge TB, et al. (2013) From green to red: Horizontal gene transfer of the phycoerythrin gene cluster between Planktothrix strains. Appl Environ Microbiol 79: 6803–6812. doi: 10.1128/AEM.01455-13
|
| [42] | Gerphagnon M, Latour D, Colombet J, et al. (2013) Fungal parasitism: life cycle, dynamics and impact on cyanobacterial blooms. PLoS One 8: 10. |
| [43] | Edvardsen B (2002) Overvåking av toksinproduserende cyanobakterier i Steinsfjorden 2001. Sammenfattende resultater fra 1997–2001. NIVA Rep 4509: 40. |
| [44] |
Su JF, Ma M, Wei L, et al. (2016) Algicidal and denitrification characterization of Acinetobacter sp J25 against Microcystis aeruginosa and microbial community in eutrophic landscape water. Mar Pollut Bull 107: 233–239. doi: 10.1016/j.marpolbul.2016.03.066
|
| [45] |
Liu YM, Wang MH, Jia RB, et al. (2012) Removal of cyanobacteria by an Aeromonas sp. Desalin Water Treat 47: 205–210. doi: 10.1080/19443994.2012.696805
|
| [46] |
Kim BR, Nakano S, Kim BH, et al. (2006) Grazing and growth of the heterotrophic flagellate Diphylleia rotans on the cyanobacterium Microcystis aeruginosa. Aquat Microb Ecol 45: 163–170. doi: 10.3354/ame045163
|
| [47] |
Zhang L, Gu L, Wei Q, et al. (2017) High temperature favors elimination of toxin-producing Microcystis and degradation of microcystins by mixotrophic Ochromonas. Chemosphere 172: 96–102. doi: 10.1016/j.chemosphere.2016.12.146
|
| [48] |
Chu TC, Murray SR, Hsu SF, et al. (2011) Temperature-induced activation of freshwater Cyanophage AS-1 prophage. Acta Histochem 113: 294–299. doi: 10.1016/j.acthis.2009.11.003
|
| [49] | Bruning K (1991) Infection of the diatom Asterionella by a chytrid. I. Effects of light on reproduction and infectivity of the parasite. J Plankton Res 13: 103–117. |
| [50] |
Visser PM, Verspagen JMH, Sandrini G, et al. (2016) How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae 54: 145–159. doi: 10.1016/j.hal.2015.12.006
|
| [51] | Rohrlack T, Haande S, Molversmyr A, et al. (2015) Environmental conditions determine the course and outcome of phytoplankton chytridiomycosis. PLoS One 10: 17. |
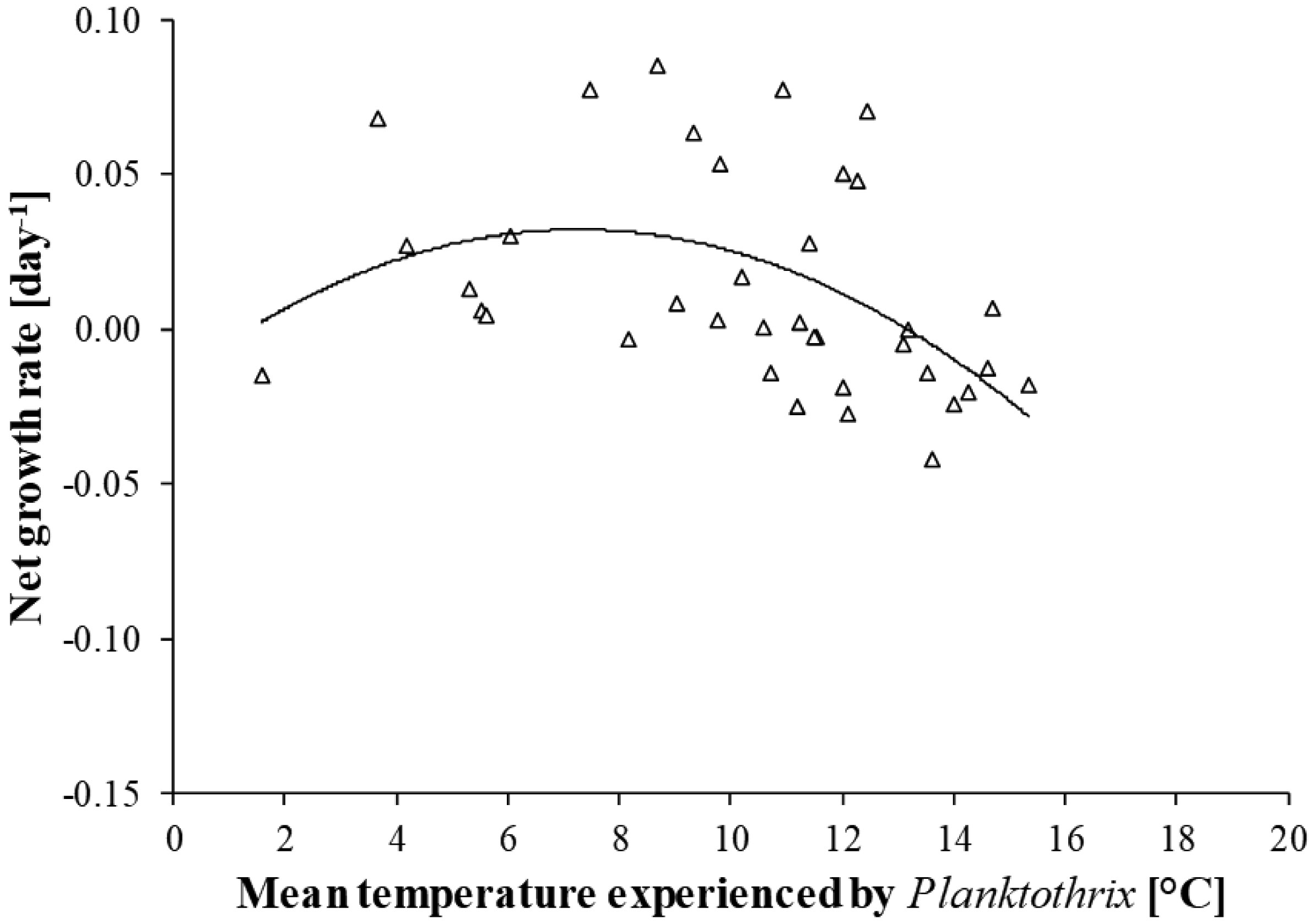
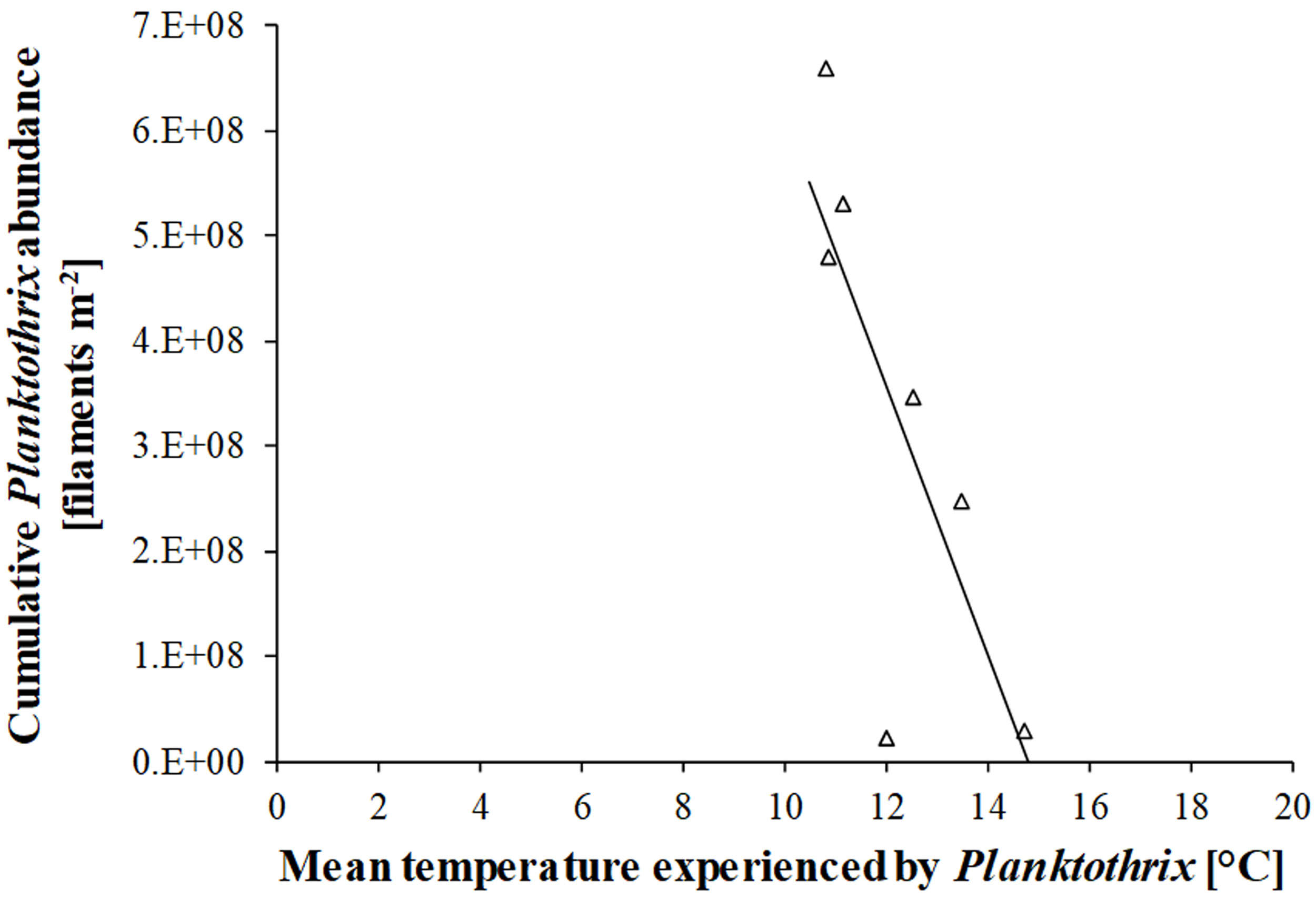
Figures(6)

Thomas Rohrlack. Low temperatures can promote cyanobacterial bloom formation by providing refuge from microbial antagonists[J]. AIMS Microbiology, 2018, 4(2): 304-318. doi: 10.3934/microbiol.2018.2.304







 DownLoad:
DownLoad: