Citation: Salma M. AlDallal. Quick glance at Fanconi anemia and BRCA2/FANCD1[J]. AIMS Medical Science, 2019, 6(4): 326-336. doi: 10.3934/medsci.2019.4.326

| [1] | Rosenberg PS, Tamary H, Alter BP (2011) How high are carrier frequencies of rare recessive syndromes? Contemporary estimates for Fanconi Anemia in the United States and Israel. Am J Med Genet A 155: 1877–1883. |
| [2] |
Kee Y, D'Andrea AD (2012) Molecular pathogenesis and clinical management of Fanconi Anemia. J Clin Invest 122: 3799–3806. doi: 10.1172/JCI58321

|
| [3] |
Romick-Rosendale LE, Hoskins EE, Privette LM, et al. (2016) Defects in the Fanconi Anemia pathway in head and neck cancer cells stimulate tumor cell invasion through DNA-PK and Rac1 signaling. Clin Cancer Res 22: 2062–2073. doi: 10.1158/1078-0432.CCR-15-2209
|
| [4] |
Romick-Rosendale LE, Lui VW, Grandis JR, et al. (2013) The Fanconi anemia pathway: Repairing the link between DNA damage and squamous cell carcinoma. Mutat Res 743: 78–88. doi: 10.1016/j.mrfmmm.2013.01.001
|
| [5] |
Shimamura A, Alter B (2010) Pathophysiology and management of inherited bone marrow failure syndromes. Blood Rev 24: 101–122. doi: 10.1016/j.blre.2010.03.002
|
| [6] |
Alan D, D'Andrea M (2010) The Fanconi anemia and breast cancer susceptibility pathways. N Engl J Med 362: 1909–1919. doi: 10.1056/NEJMra0809889
|
| [7] |
Taniguchi T, D'Andrea A (2006) Molecular pathogenesis of Fanconi anemia: recent progress. Blood 107: 4223–4233. doi: 10.1182/blood-2005-10-4240
|
| [8] |
Nepal M, Che R, Ma C, et al. (2017) FANCD2 and DNA damage. Int J Mol Sci 18: 1804. doi: 10.3390/ijms18081804
|
| [9] |
Che R, Zhang J, Nepal M, et al. (2018) Multifaceted Fanconi anemia signaling. Trends Genet 34: 171–183. doi: 10.1016/j.tig.2017.11.006
|
| [10] |
Strathdee CA, Gavish H, Shannon WR, et al. (1992) Cloning of cDNAs for Fanconi's anaemia by functional complementation. Nature 356: 763–767. doi: 10.1038/356763a0
|
| [11] | Chen H, Zhan S, Wu Z (2014) Fanconi anemia pathway defects in inherited and sporadic cancers. Transl Pediatr 3: 300–304. |
| [12] |
Cancer Genome Atlas Research Network (2011) Integrated genomic analyses of ovarian carcinoma. Nature 474: 609–615. doi: 10.1038/nature10166
|
| [13] |
Sumpter R, Levine B (2017) Emerging functions of the Fanconi anemia pathway at a glance. J Cell Sci 130: 2657–2662. doi: 10.1242/jcs.204909
|
| [14] |
Yamashita T, Wu N, Kupfer G, et al. (1996) Clinical variability of Fanconi anemia (type C) results from expression of an amino terminal truncated Fanconi anemia complementation group C polypeptide with partial activity. Blood 87: 4424–4432. doi: 10.1182/blood.V87.10.4424.bloodjournal87104424
|
| [15] |
Whitney MA, Royle G, Low MJ, et al. (1996) Germ cell defects and hematopoietic hypersensitivity to gamma-interferon in mice with a targeted disruption of the Fanconi anemia C gene. Blood 88: 49–58. doi: 10.1182/blood.V88.1.49.49
|
| [16] |
Richardson C, Yan S, Vestal CG (2015) Oxidative stress, bone marrow failure, and genome instability in hematopoietic stem cells. Int J Mol Sci 16: 2366–2385. doi: 10.3390/ijms16022366
|
| [17] |
Smogorzewska A, Matsuoka S, Vinciguerra P, et al. (2007) Identification of the FANCI protein, a monoubiquitinated FANCD2 paralog required for DNA repair. Cell 129: 289–301. doi: 10.1016/j.cell.2007.03.009
|
| [18] |
Garcia-Higuera I, Taniguchi T, Ganesan S, et al. (2001) Interaction of the Fanconi anemia proteins and BRCA1 in a common pathway. Mol Cell 7: 249–262. doi: 10.1016/S1097-2765(01)00173-3
|
| [19] |
Sims AE, Spiteri E, Sims RJ 3rd, et al. (2007) FANCI is a second monoubiquitinated member of the Fanconi anemia pathway. Nat Struct Mol Biol 14: 564–567. doi: 10.1038/nsmb1252
|
| [20] | Dorsman JC, Levitus M, Rockx D, et al. (2007) Identification of the Fanconianemiacomplementation group I gene, FANCI. Cell Oncol 29: 211–218. |
| [21] |
Kratz K, Schöpf B, Kaden S, et al. (2010) Deficiency of FANCD2-associated nuclease KIAA1018/ FAN1 sensitizes cells to interstrand crosslinking agents. Cell 142: 77–88. doi: 10.1016/j.cell.2010.06.022
|
| [22] |
Liu T, Ghosal G, Yuan J, et al. (2010) FAN1 acts with FANCI-FANCD2 to promote DNA interstrand cross-link repair. Science 329: 693–696. doi: 10.1126/science.1192656
|
| [23] |
MacKay C, Déclais AC, Lundin C, et al. (2010) Identification of KIAA1018/FAN1, a DNA repair nuclease recruited to DNA damage by monoubiquitinated FANCD2. Cell 142: 65–76. doi: 10.1016/j.cell.2010.06.021
|
| [24] |
Smogorzewska A, Desetty R, Saito TT, et al. (2010) A genetic screen identifies FAN1, a Fanconi anemia-associated nuclease necessary for DNA interstrand crosslink repair. Mol Cell 39: 36–47. doi: 10.1016/j.molcel.2010.06.023
|
| [25] |
Howlett NG, Taniguchi T, Olson S, et al. (2002) Biallelic inactivation of BRCA2 in Fanconi anemia. Science 297: 606–609. doi: 10.1126/science.1073834
|
| [26] |
Xia B, Dorsman JC, Ameziane N, et al. (2007) Fanconi anemia is associated with a defect in the BRCA2 partner PALB2. Nat Genet 39: 159–161. doi: 10.1038/ng1942
|
| [27] |
Reid S, Schindler D, Hanenberg H, et al. (2007) Biallelic mutations in PALB2 cause Fanconi anemia subtype FA-N and predispose to childhood cancer. Nat Genet 39: 162–164. doi: 10.1038/ng1947
|
| [28] |
Litman R, Peng M, Jin Z, et al. (2004) BACH1 is critical for homologous recombination and appears to be the Fanconi anemia gene product FANCJ. Cancer Cell 8: 255–265. doi: 10.1016/j.ccr.2005.08.004
|
| [29] |
Levran O, Attwooll C, Henry RT, et al. (2005) The BRCA1-interacting helicase BRIP1 is deficient in Fanconi anemia. Nat Genet 37: 931–933. doi: 10.1038/ng1624
|
| [30] |
Levitus M, Waisfisz Q, Godthelp BC, et al. (2005) The DNA helicase BRIP1 is defective in Fanconi anemia complementation group. Nat Genet 37: 934–935. doi: 10.1038/ng1625
|
| [31] |
Bridge WL, Vandenberg CJ, Franklin RJ, et al. (2005) The BRIP1 helicase functions independently of BRCA1 in the Fanconi anemia pathway for DNA crosslink repair. Nat Genet 37: 953–957. doi: 10.1038/ng1627
|
| [32] |
Crossan GP, van der Weyden L, Rosado IV, et al. (2011) Disruption of mouse Slx4, a regulator of structure specific nucleases, phenocopies Fanconi anemia. Nat Genet 43: 147–152. doi: 10.1038/ng.752
|
| [33] |
Kim Y, Lach FP, Desetty R, et al. (2011) Mutations of the SLX4 gene in Fanconi anemia. Nat Genet 43: 142–146. doi: 10.1038/ng.750
|
| [34] |
Stoepker C, Hain K, Schuster B, et al. (2011) SLX4, a coordinator of structure-specific endonucleases, is mutated in a new Fanconi anemia subtype. Nat Genet 43: 138–141. doi: 10.1038/ng.751
|
| [35] |
Fekairi S, Scaglione S, Chahwan C, et al. (2009) Human SLX4 is a Holliday junction resolvase subunit that binds multiple DNA repair/recombination endonucleases. Cell 138: 78–89. doi: 10.1016/j.cell.2009.06.029
|
| [36] |
Svendsen JM, Smogorzewska A, Sowa ME, et al. (2009) Mammalian BTBD12/SLX4 assembles a Holliday junction resolvase and is required for DNA repair. Cell 138: 63–77. doi: 10.1016/j.cell.2009.06.030
|
| [37] |
Munoz IM, Hain K, Déclais AC, et al. (2009) Coordination of structure-specific nucleases by human SLX4/BTBD12 is required for DNA repair. Mol Cell 35: 116–127. doi: 10.1016/j.molcel.2009.06.020
|
| [38] |
Kuraoka I, Kobertz WR, Ariza RR, et al. (2000) Repair of an interstrand DNA cross-link initiated by ERCC1-XPF repair/recombination nuclease. J Biol Chem 275: 26632–26636. doi: 10.1074/jbc.C000337200
|
| [39] |
Hanada K, Budzowska M, Modesti M, et al. (2006) The structure-specific endonuclease Mus81-Eme1 promotes conversion of interstrand DNA crosslinks into double-strandsbreaks. EMBO J 25: 4921–4932. doi: 10.1038/sj.emboj.7601344
|
| [40] |
Duncan JA, Reeves JR, Cooke TG (1998) BRCA1 and BRCA2 proteins: roles in health and disease. Mol Pathol 51: 237–247. doi: 10.1136/mp.51.5.237
|
| [41] |
Yoshida K, Miki Y (2004) Role of BRCA1 and BRCA2 as regulators of DNA repair, transcription, and cell cycle in response to DNA damage. Cancer Sci 95: 866–871. doi: 10.1111/j.1349-7006.2004.tb02195.x
|
| [42] |
Mijic S, ZellwegerR, Chappidi N, et al. (2017) Replication fork reversal triggers fork degradation in BRCA2-defective cells. Nat Commun 8: 859–859. doi: 10.1038/s41467-017-01164-5
|
| [43] |
O'Donovan PJ, Livingston DM (2010) BRCA1 and BRCA2: breast/ovarian cancer susceptibility gene products and participants in DNA double-strand break repair. Carcinogenesis 31: 961–967. doi: 10.1093/carcin/bgq069
|
| [44] |
Alter B, Rosenberg P, Brody L (2007) Clinical and molecular features associated with biallelic mutations in FANCD1/BRCA2. J Med Genet 44: 1–9. doi: 10.1136/jmg.2006.043257
|
| [45] |
García MJ, Fernández V, Osorio A, et al. (2009) Mutational analysis of FANCL, FANCM and the recently identified FANCI suggests that among the 13 known Fanconi Anemia genes, only FANCD1/BRCA2 plays a major role in high-risk breast cancer predisposition. Carcinogenesis 30: 1898–1902. doi: 10.1093/carcin/bgp218
|
| [46] |
Myers K, Davies S, Harris R, et al. (2012) The clinical phenotype of children with fanconi anemia caused by biallelic FANCD1/BRCA2 mutations. Pediatr Blood Cancer 58: 462–465. doi: 10.1002/pbc.23168
|
| [47] |
Petrucelli N, Daly M, Feldman G. (2010) Hereditary breast and ovarian cancer due to mutations in BRCA1 and BRCA2. Genet Med 12: 245–259. doi: 10.1097/GIM.0b013e3181d38f2f
|
| [48] |
Bakker S, Vrugt HV, Rooimans M, et al. (2009) Fancm-deficient mice reveal unique features of Fanconi anemia complementation group M. Human Mol Genet 18: 3484–3495. doi: 10.1093/hmg/ddp297
|
| [49] |
Dong H, Nebert D, Bruford E, et al. (2015) Update of the human and mouse Fanconi anemia genes. Hum Genomics 9: 2–10. doi: 10.1186/s40246-015-0024-4
|
| [50] |
Hirsch B, Shimamura A, Moreau L, et al. (2004) Association of biallelic BRCA2/FANCD1 mutations with spontaneous chromosomal instability and solid tumors of childhood. Blood 103: 2554–2559. doi: 10.1182/blood-2003-06-1970
|
| [51] |
Hahn S, Greenhalf B, Ellis I, et al. (2003) BRCA2 germline mutations in familial pancreatic carcinoma. J Natl Cancer Inst 95: 214–221. doi: 10.1093/jnci/95.3.214
|
| [52] |
Liede A, Karlan B, Narod S (2004) Cancer risks for male carriers of germline mutations in BRCA1 or BRCA2: A review of the literature. J Clin Oncol 22: 735–742. doi: 10.1200/jco.2004.22.14_suppl.735
|
| [53] |
Degrolard-Courcet E, Sokolowska J, Padeano MM, et al. (2014) Development of primary early-onset colorectal cancers due to biallelic mutations of the FANCD1/BRCA2 gene. Eur J Hum Genet 22: 979–987. doi: 10.1038/ejhg.2013.278
|
| [54] |
Akbari MR, Malekzadeh R, Lepage P, et al. (2011) Mutations in Fanconi anemia genes and the risk of esophageal cancer. Hum Genet 129: 573–582. doi: 10.1007/s00439-011-0951-7
|
| [55] |
Dodgshun AJ, Sexton-Oates A, Saffery R, et al. (2016) Biallelic FANCD1/BRCA2 mutations predisposing to glioblastoma multiforme with multiple oncogenic amplifications. Cancer Genet 209: 53–56. doi: 10.1016/j.cancergen.2015.11.005
|
| [56] |
Dewire MD, Ellison DW, Patay Z, et al. (2009) Fanconi anemia and biallelic BRCA2 mutation diagnosed in a young child with an embryonal CNS tumor. Pediatr Blood Cancer 53: 1140–1142. doi: 10.1002/pbc.22139
|
| [57] |
Elledge S, Amon A (2002) The BRCA1 suppressor hypothesis: an explanation for the tissue- specific tumor development in BRCA1 patients. Cancer Cell 1: 129–132. doi: 10.1016/S1535-6108(02)00041-7
|
| [58] | Ford D, Easton DF, Bishop DT, et al. (1994) Risks of cancer in BRCA1-mutation carrieirs. Breast cancer linkage consortium. Lancet 343: 692–695. |
| [59] |
Berwick M, Satagopan J, Ben-Porat L, et al. (2007) Genetic heterogeneity among Fanconi anemia heterozygotes and risk of cancer. Cancer Res 67: 9591–9596. doi: 10.1158/0008-5472.CAN-07-1501
|
| [60] | Skvarova KK, Osborn MJ, Webber BR, et al. (2017) CRISPR/Cas9-Mediated Correction of the FANCD1 Gene in Primary Patient Cells. J Mol Med 18: 1269–1269. |
| [61] |
Chen C, Kennedy R, Sidi S (2009) Chk1 inhibition as a strategy for targeting Fanconi anemia (FA) DNA repair pathway defiecient tumors. Mol Cancer 8: 1–24. doi: 10.1186/1476-4598-8-24
|
| [62] |
Bryant H, Helleday T (2006) Inhibition of poly (ADP-ribose) polymerase activates ATM which is required for subsequent homologous recombination repair. Nucleic Acids Res 34: 1685–1691. doi: 10.1093/nar/gkl108
|
| [63] |
Farmer H, McCabe N, Lord CJ, et al. (2005) Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature 434: 917–921. doi: 10.1038/nature03445
|
| [64] |
Fong PC, Boss DS, Yap TA, et al. (2009) Inhibition of poly (ADP-ribose) polymerase in tumor from BRCA mutation carriers. N Engl J Med 361: 123–134. doi: 10.1056/NEJMoa0900212
|
| [65] |
Chen C, Taniguchi T, D'Andrea A (2007) The Fanconi anemia pathway confers glioma resistance to DNA alkylating agents. J Mol Med 85: 497–509. doi: 10.1007/s00109-006-0153-2
|
| [66] |
Levran O, Attwooll C, Henry RT, et al. (2005) The BRCA1-interacting helicase BRIP1 is deficient in Fanconi anemia. Nat Genet 37: 931–933. doi: 10.1038/ng1624
|
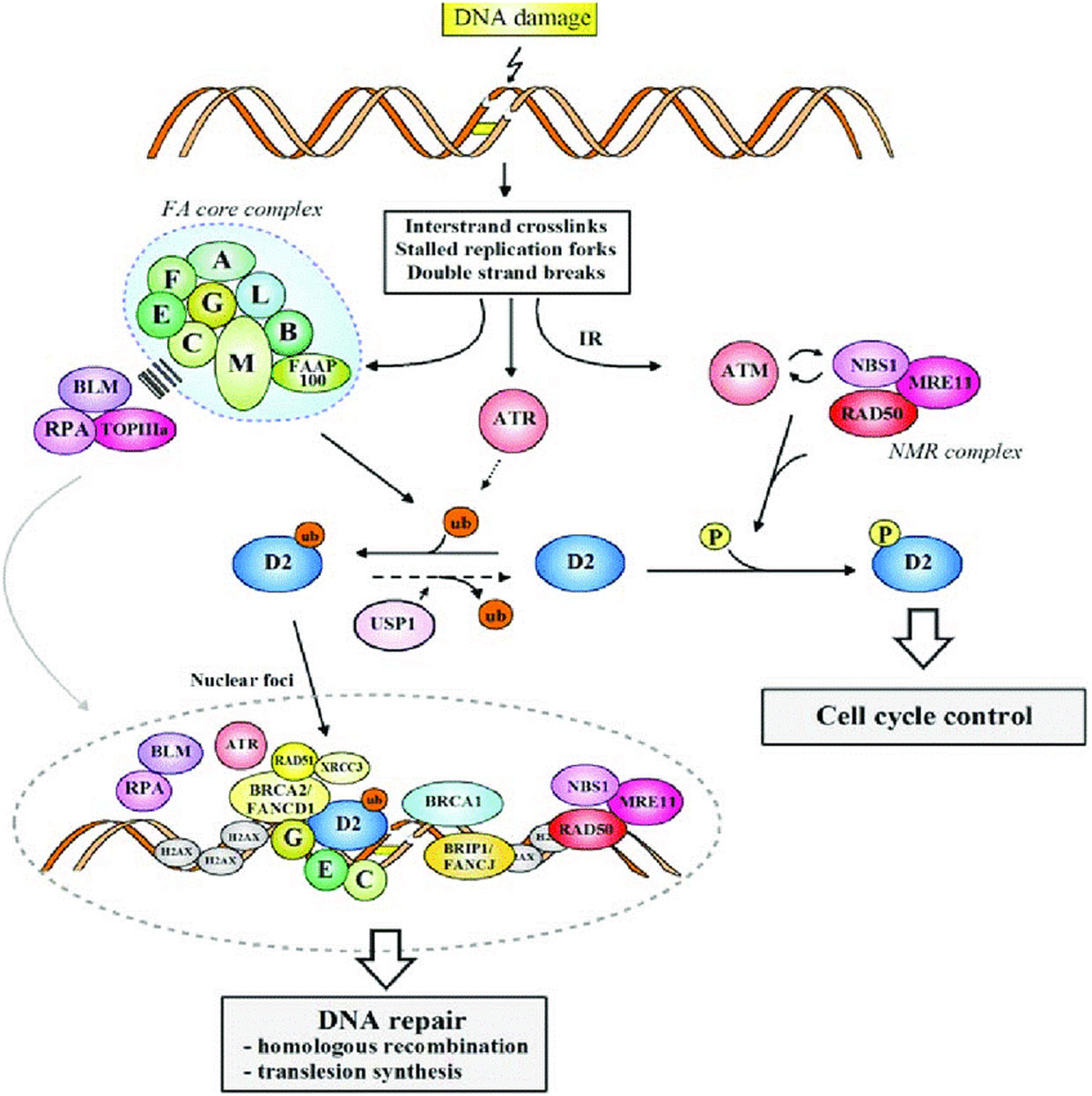
Figures(1) / Tables(1)

Salma M. AlDallal. Quick glance at Fanconi anemia and BRCA2/FANCD1[J]. AIMS Medical Science, 2019, 6(4): 326-336. doi: 10.3934/medsci.2019.4.326







 DownLoad:
DownLoad: