Citation: Roberto Gutierrez, Yesenia Thompson, Timothy R. O’Connor. DNA direct repair pathways in cancer[J]. AIMS Medical Science, 2018, 5(3): 284-302. doi: 10.3934/medsci.2018.3.284

| [1] |
De Bont R, van Larebeke N (2004) Endogenous DNA damage in humans: A review of quantitative data. Mutagenesis 19: 169–185. doi: 10.1093/mutage/geh025

|
| [2] | Rydberg B, Lindahl T (1982) Nonenzymatic methylation of DNA by the intracellular methyl group donor S-adenosyl-L-methionine is a potentially mutagenic reaction. EMBO J 1: 211–216. |
| [3] |
Ballschmiter K (2003) Pattern and sources of naturally produced organohalogens in the marine environment: Biogenic formation of organohalogens. Chemosphere 52: 313–324. doi: 10.1016/S0045-6535(03)00211-X
|
| [4] |
Hamilton JT, Mcroberts WC, Keppler F, et al. (2003) Chloride methylation by plant pectin: An efficient environmentally significant process. Science 301: 206–209. doi: 10.1126/science.1085036
|
| [5] |
Fu D, Calvo JA, Samson LD (2012) Balancing repair and tolerance of DNA damage caused by alkylating agents. Nat Rev Cancer 12: 104–120. doi: 10.1038/nrc3185
|
| [6] |
Singer B (1975) The chemical effects of nucleic acid alkylation and their relation to mutagenesis and carcinogenesis. Prog Nucleic Acid Res Mol Biol 15: 219–284. doi: 10.1016/S0079-6603(08)60121-X
|
| [7] | Singer B, Grunberger D (1983) Reaction of directly acting agents with nucleic acids, In: Singer B, Grunberger D. Molecular biology of mutagens and carcinogens, Boston: Springer, 45–96. |
| [8] |
Beranek DT (1990) Distribution of methyl and ethyl adducts following alkylation with monofunctional alkylating agents. Mutat Res 231: 11–30. doi: 10.1016/0027-5107(90)90173-2
|
| [9] |
Sukumar S, Notario V, Martin-Zanca D, et al. (1983) Induction of mammary carcinomas in rats by nitroso-methylurea involves malignant activation of H-ras-1 locus by single point mutations. Nature 306: 658–661. doi: 10.1038/306658a0
|
| [10] |
Zarbl H, Sukumar S, Arthur AV, et al. (1985) Direct mutagenesis of Ha-ras-1 oncogenes by N-nitroso-N-methylurea during initiation of mammary carcinogenesis in rats. Nature 315: 382–385. doi: 10.1038/315382a0
|
| [11] |
Esteller M, Garcia-Foncillas J, Andion E, et al. (2000) Inactivation of the DNA-repair gene MGMT and the clinical response of gliomas to alkylating agents. N Engl J Med 343: 1350–1354. doi: 10.1056/NEJM200011093431901
|
| [12] | Christmann M, Kaina B (2016) MGMT-a critical DNA repair gene target for chemotherapy resistance, In: Kelley MR, Fischel ML. DNA repair in cancer therapy, New York: Elsevier, 55–82. |
| [13] | Ahmad A, Nay SL, O'Connor TR (2015) Direct Reversal Repair in Mammalian Cells, In: Chen C. Advances in DNA Repair: IntechOpen, 95–128. |
| [14] | Lindahl T, Demple B, Robins P (1982) Suicide inactivation of the E. coli O6-methylguanine-DNA methyltransferase. EMBO J 1: 1359–1363. |
| [15] |
Loenarz C, Schofield CJ (2011) Physiological and biochemical aspects of hydroxylations and demethylations catalyzed by human 2-oxoglutarate oxygenases. Trends Biochem Sci 36: 7–18. doi: 10.1016/j.tibs.2010.07.002
|
| [16] |
Knijnenburg TA, Wang L, Zimmermann MT, et al. (2018) Genomic and Molecular Landscape of DNA Damage Repair Deficiency across The Cancer Genome Atlas. Cell Rep 23: 239–254. doi: 10.1016/j.celrep.2018.03.076
|
| [17] |
Wiewrodt D, Nagel G, Dreimuller N, et al. (2008) MGMT in primary and recurrent human glioblastomas after radiation and chemotherapy and comparison with p53 status and clinical outcome. Int J Cancer 122: 1391–1399. doi: 10.1002/ijc.23219
|
| [18] |
Gao W, Li L, Xu P, et al. (2011) Frequent down-regulation of hABH2 in gastric cancer and its involvement in growth of cancer cells. J Gastroenterol Hepatol 26: 577–584. doi: 10.1111/j.1440-1746.2010.06531.x
|
| [19] |
Tasaki M, Shimada K, Kimura H, et al. (2011) ALKBH3, a human AlkB homologue, contributes to cell survival in human non-small-cell lung cancer. Br J Cancer 104: 700–706. doi: 10.1038/sj.bjc.6606012
|
| [20] |
Yamato I, Sho M, Shimada K, et al. (2012) PCA-1/ALKBH3 contributes to pancreatic cancer by supporting apoptotic resistance and angiogenesis. Cancer Res 72: 4829–4839. doi: 10.1158/0008-5472.CAN-12-0328
|
| [21] |
Hegi ME, Diserens AC, Gorlia T, et al. (2005) MGMT gene silencing and benefit from temozolomide in glioblastoma. N Engl J Med 352: 997–1003. doi: 10.1056/NEJMoa043331
|
| [22] |
Hermisson M, Klumpp A, Wick W, et al. (2006) O6-methylguanine DNA methyltransferase and p53 status predict temozolomide sensitivity in human malignant glioma cells. J Neurochem 96: 766–776. doi: 10.1111/j.1471-4159.2005.03583.x
|
| [23] |
Soll JM, Sobol RW, Mosammaparast N (2017) Regulation of DNA alkylation damage repair: Lessons and therapeutic opportunities. Trends Biochem Sci 42: 206–218. doi: 10.1016/j.tibs.2016.10.001
|
| [24] |
Weller M, Tabatabai G, Kastner B, et al. (2015) MGMT promoter methylation is a strong prognostic biomarker for benefit from dose-intensified temozolomide rechallenge in progressive glioblastoma: The DIRECTOR Trial. Clin Cancer Res 21: 2057–2064. doi: 10.1158/1078-0432.CCR-14-2737
|
| [25] |
Verbeek B, Southgate TD, Gilham DE, et al. (2008) O6-Methylguanine-DNA methyltransferase inactivation and chemotherapy. Br Med Bull 85: 17–33. doi: 10.1093/bmb/ldm036
|
| [26] |
Tubbs JL, Pegg AE, Tainer JA (2007) DNA binding, nucleotide flipping, and the helix-turn-helix motif in base repair by O6-alkylguanine-DNA alkyltransferase and its implications for cancer chemotherapy. DNA Repair 6: 1100–1115. doi: 10.1016/j.dnarep.2007.03.011
|
| [27] |
Duguid EM, Rice PA, He C (2005) The structure of the human AGT protein bound to DNA and its implications for damage detection. J Mol Biol 350: 657–666. doi: 10.1016/j.jmb.2005.05.028
|
| [28] |
Daniels DS, Tainer JA (2000) Conserved structural motifs governing the stoichiometric repair of alkylated DNA by O6-alkylguanine-DNA alkyltransferase. Mutat Res 460: 151–163. doi: 10.1016/S0921-8777(00)00024-0
|
| [29] |
Pegg AE (2000) Repair of O6-alkylguanine by alkyltransferases. Mutat Res 462: 83–100. doi: 10.1016/S1383-5742(00)00017-X
|
| [30] | Demple B, Jacobsson A, Olsson M, et al. (1982) Repair of alkylated DNA in Escherichia coli. Physical properties of O6-methylguanine-DNA methyltransferase. J Biol Chem 257: 13776–13780. |
| [31] | Liu L, Xu-Welliver M, Kanugula S, et al. (2002) Inactivation and degradation of O6-alkylguanine-DNA alkyltransferase after reaction with nitric oxide. Cancer Res 62: 3037–3043. |
| [32] |
Xu-Welliver M, Pegg AE (2002) Degradation of the alkylated form of the DNA repair protein, O6-alkylguanine-DNA alkyltransferase. Carcinogenesis 23: 823–830. doi: 10.1093/carcin/23.5.823
|
| [33] |
Rasimas JJ, Dalessio PA, Ropson IJ, et al. (2004) Active-site alkylation destabilizes human O6-alkylguanine DNA alkyltransferase. Protein Sci 13: 301–305. doi: 10.1110/ps.03319404
|
| [34] |
Oh HK, Teo AK, Ali RB, et al. (1996) Conformational change in human DNA repair enzyme O6-methylguanine-DNA methyltransferase upon alkylation of its active site by SN1 (indirect-acting) and SN2 (direct-acting) alkylating agents: Breaking a "salt-link". Biochemistry 35: 12259–12266. doi: 10.1021/bi9603635
|
| [35] |
Lindahl T, Barnes DE (1992) Mammalian DNA ligases. Annu Rev Biochem 61: 251–281. doi: 10.1146/annurev.bi.61.070192.001343
|
| [36] | Lefebvre P, Laval F (1986) Enhancement of O6-methylguanine-DNA-methyltransferase activity induced by various treatments in mammalian cells. Cancer Res 46: 5701–5705. |
| [37] |
Frosina G, Laval F (1987) The O6-methylguanine-DNA-methyltransferase activity of rat hepatoma cells is increased after a single exposure to alkylating agents. Carcinogenesis 8: 91–95. doi: 10.1093/carcin/8.1.91
|
| [38] |
Laval F (1990) Induction of proteins involved in the repair of alkylated bases in mammalian cells by DNA-damaging agents. Mutat Res 233: 211–218. doi: 10.1016/0027-5107(90)90164-Y
|
| [39] |
Aasland D, Reich TR, Tomicic MT, et al. (2018) Repair gene O6-methylguanine-DNA methyltransferase is controlled by SP1 and up-regulated by glucocorticoids, but not by temozolomide and radiation. J Neurochem 144: 139–151. doi: 10.1111/jnc.14262
|
| [40] |
Kreth S, Limbeck E, Hinske LC, et al. (2013) In human glioblastomas transcript elongation by alternative polyadenylation and miRNA targeting is a potent mechanism of MGMT silencing. Acta Neuropathol 125: 671–681. doi: 10.1007/s00401-013-1081-1
|
| [41] | Ramakrishnan V, Kushwaha D, Koay DC, et al. (2011) Post-transcriptional regulation of O6-methylguanine-DNA methyltransferase MGMT in glioblastomas. Cancer Biomark 10: 185–193. |
| [42] |
Zhang W, Zhang J, Hoadley K, et al. (2012) miR-181d: A predictive glioblastoma biomarker that downregulates MGMT expression. Neuro Oncol 14: 712–719. doi: 10.1093/neuonc/nos089
|
| [43] |
Gao YT, Chen XB, Liu HL (2016) Up-regulation of miR-370-3p restores glioblastoma multiforme sensitivity to temozolomide by influencing MGMT expression. Sci Rep 6: 32972. doi: 10.1038/srep32972
|
| [44] |
Swann PF, Magee PN (1969) Induction of rat kidney tumours by ethyl methanesulphonate and nervous tissue tumours by methyl methanesulphonate and ethyl methanesulphonate. Nature 223: 947–949. doi: 10.1038/223947a0
|
| [45] | Snow ET, Foote RS, Mitra S (1984) Base-pairing properties of O6-methylguanine in template DNA during in vitro DNA replication. J Biol Chem 259: 8095–8100. |
| [46] |
Van Houten B, Sancar A (1987) Repair of N-methyl-N'-nitro-N-nitrosoguanidine-induced DNA damage by ABC excinuclease. J Bacteriol 169: 540–545. doi: 10.1128/jb.169.2.540-545.1987
|
| [47] |
Glassner BJ, Weeda G, Allan JM, et al. (1999) DNA repair methyltransferase (Mgmt) knockout mice are sensitive to the lethal effects of chemotherapeutic alkylating agents. Mutagenesis 14: 339–347. doi: 10.1093/mutage/14.3.339
|
| [48] |
Toorchen D, Topal MD (1983) Mechanisms of chemical mutagenesis and carcinogenesis: Effects on DNA replication of methylation at the O6-guanine position of dGTP. Carcinogenesis 4: 1591–1597. doi: 10.1093/carcin/4.12.1591
|
| [49] |
Choi JY, Chowdhury G, Zang H, et al. (2006) Translesion synthesis across O6-alkylguanine DNA adducts by recombinant human DNA polymerases. J Biol Chem 281: 38244–38256. doi: 10.1074/jbc.M608369200
|
| [50] |
Delaney JC, Essigmann JM (2001) Effect of Sequence Context on O6-Methylguanine Repair and Replication in vivo. Biochemistry 40: 14968–14975. doi: 10.1021/bi015578f
|
| [51] |
Pence MG, Choi JY, Egli M, et al. (2010) Structural basis for proficient incorporation of dTTP opposite O6-methylguanine by human DNA polymerase iota. J Biol Chem 285: 40666–40672. doi: 10.1074/jbc.M110.183665
|
| [52] |
Day RS, Ziolkowski CH, Scudiero DA, et al. (1980) Defective repair of alkylated DNA by human tumour and SV40-transformed human cell strains. Nature 288: 724–727. doi: 10.1038/288724a0
|
| [53] | Hansen RJ, Nagasubramanian R, Delaney SM, et al. (2007) Role of O6-methylguanine-DNA methyltransferase in protecting from alkylating agent-induced toxicity and mutations in mice. Carcinogenesis 28: 1111–1116. |
| [54] |
Kaina B, Christmann M, Naumann S, et al. (2007) MGMT: Key node in the battle against genotoxicity, carcinogenicity and apoptosis induced by alkylating agents. DNA Repair 6: 1079–1099. doi: 10.1016/j.dnarep.2007.03.008
|
| [55] | Sakumi K, Shiraishi A, Shimizu S, et al. (1997) Methylnitrosourea-induced tumorigenesis in MGMT gene knockout mice. Cancer Res 57: 2415–2418. |
| [56] |
Tsuzuki T, Sakumi K, Shiraishi A, et al. (1996) Targeted disruption of the DNA repair methyltransferase gene renders mice hypersensitive to alkylating agent. Carcinogenesis 17: 1215–1220. doi: 10.1093/carcin/17.6.1215
|
| [57] |
Reese JS, Allay E, Gerson SL (2001) Overexpression of human O6-alkylguanine DNA alkyltransferase (AGT) prevents MNU induced lymphomas in heterozygous p53 deficient mice. Oncogene 20: 5258–5263. doi: 10.1038/sj.onc.1204700
|
| [58] |
Wirtz S, Nagel G, Eshkind L, et al. (2010) Both base excision repair and O6-methylguanine-DNA methyltransferase protect against methylation-induced colon carcinogenesis. Carcinogenesis 31: 2111–2117. doi: 10.1093/carcin/bgq174
|
| [59] |
Psaroudi MC, Kyrtopoulos SA (2000) Toxicity, mutation frequency and mutation spectrum induced by dacarbazine in CHO cells expressing different levels of O6-methylguanine-DNA methyltransferase. Mutat Res 447: 257–265. doi: 10.1016/S0027-5107(99)00220-1
|
| [60] |
Quiros S, Roos WP, Kaina B (2010) Processing of O6-methylguanine into DNA double-strand breaks requires two rounds of replication whereas apoptosis is also induced in subsequent cell cycles. Cell Cycle 9: 168–178. doi: 10.4161/cc.9.1.10363
|
| [61] |
Mojas N, Lopes M, Jiricny J (2007) Mismatch repair-dependent processing of methylation damage gives rise to persistent single-stranded gaps in newly replicated DNA. Genes Dev 21: 3342–3355. doi: 10.1101/gad.455407
|
| [62] |
York SJ, Modrich P (2006) Mismatch repair-dependent iterative excision at irreparable O6-methylguanine lesions in human nuclear extracts. J Biol Chem 281: 22674–22683. doi: 10.1074/jbc.M603667200
|
| [63] | Ochs K, Kaina B (2000) Apoptosis induced by DNA damage O6-methylguanine is Bcl-2 and caspase-9/3 regulated and Fas/caspase-8 independent. Cancer Res 60: 5815–5824. |
| [64] |
Hunter C, Smith R, Cahill DP, et al. (2006) A hypermutation phenotype and somatic MSH6 mutations in recurrent human malignant gliomas after alkylator chemotherapy. Cancer Res 66: 3987–3991. doi: 10.1158/0008-5472.CAN-06-0127
|
| [65] |
Patel SA, Graunke DM, Pieper RO (1997) Aberrant silencing of the CpG island-containing human O6-methylguanine DNA methyltransferase gene is associated with the loss of nucleosome-like positioning. Mol Cell Biol 17: 5813–5822. doi: 10.1128/MCB.17.10.5813
|
| [66] |
Watts GS, Pieper RO, Costello JF, et al. (1997) Methylation of discrete regions of the O6-methylguanine DNA methyltransferase (MGMT) CpG island is associated with heterochromatinization of the MGMT transcription start site and silencing of the gene. Mol Cell Biol 17: 5612–5619. doi: 10.1128/MCB.17.9.5612
|
| [67] |
Kaina B, Fritz G, Mitra S, et al. (1991) Transfection and expression of human O6-methylguanine-DNA methyltransferase (MGMT) cDNA in Chinese hamster cells: The role of MGMT in protection against the genotoxic effects of alkylating agents. Carcinogenesis 12: 1857–1867. doi: 10.1093/carcin/12.10.1857
|
| [68] |
Dumenco LL, Allay E, Norton K, et al. (1993) The prevention of thymic lymphomas in transgenic mice by human O6-alkylguanine-DNA alkyltransferase. Science 259: 219–222. doi: 10.1126/science.8421782
|
| [69] | Liu L, Allay E, Dumenco LL, et al. (1994) Rapid repair of O6-methylguanine-DNA adducts protects transgenic mice from N-methylnitrosourea-induced thymic lymphomas. Cancer Res 54: 4648–4652. |
| [70] |
Zhou ZQ, Manguino D, Kewitt K, et al. (2001) Spontaneous hepatocellular carcinoma is reduced in transgenic mice overexpressing human O6-methylguanine-DNA methyltransferase. Proc Natl Acad Sci U S A 98: 12566–12571. doi: 10.1073/pnas.221232998
|
| [71] |
Zaidi NH, Pretlow TP, O'Riordan MA, et al. (1995) Transgenic expression of human MGMT protects against azoxymethane-induced aberrant crypt foci and G to A mutations in the K-ras oncogene of mouse colon. Carcinogenesis 16: 451–456. doi: 10.1093/carcin/16.3.451
|
| [72] |
Nakatsuru Y, Matsukuma S, Nemoto N, et al. (1993) O6-methylguanine-DNA methyltransferase protects against nitrosamine-induced hepatocarcinogenesis. Proc Natl Acad Sci U S A 90: 6468–6472. doi: 10.1073/pnas.90.14.6468
|
| [73] |
Allay E, Veigl M, Gerson SL (1999) Mice over-expressing human O6-alkylguanine-DNA alkyltransferase selectively reduce O6-methylguanine mediated carcinogenic mutations to threshold levels after N-methyl-N-nitrosourea. Oncogene 18: 3783–3787. doi: 10.1038/sj.onc.1202697
|
| [74] |
Allay E, Reese JS, Mcguire EA, et al. (1997) Potentiation of lymphomagenesis by methylnitrosourea in mice transgenic for LMO1 is blocked by O6-alkylguanine DNA-alkyltransferase. Oncogene 15: 2127–2132. doi: 10.1038/sj.onc.1201384
|
| [75] |
Qin X, Zhou H, Liu L, et al. (1999) Transgenic expression of human MGMT blocks the hypersensitivity of PMS2-deficient mice to low dose MNU thymic lymphomagenesis. Carcinogenesis 20: 1667–1673. doi: 10.1093/carcin/20.9.1667
|
| [76] | Becker K, Dosch J, Gregel CM, et al. (1996) Targeted expression of human O6-methylguanine-DNA methyltransferase (MGMT) in transgenic mice protects against tumor initiation in two-stage skin carcinogenesis. Cancer Res 56: 3244–3249. |
| [77] |
Becker K, Gregel C, Fricke C, et al. (2003) DNA repair protein MGMT protects against N-methyl-N-nitrosourea-induced conversion of benign into malignant tumors. Carcinogenesis 24: 541–546. doi: 10.1093/carcin/24.3.541
|
| [78] |
Kurowski MA, Bhagwat AS, Papaj G, et al. (2003) Phylogenomic identification of five new human homologs of the DNA repair enzyme AlkB. BMC Genomics 4: 48. doi: 10.1186/1471-2164-4-48
|
| [79] |
Gerken T, Girard CA, Tung YC, et al. (2007) The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase. Science 318: 1469–1472. doi: 10.1126/science.1151710
|
| [80] |
Duncan T, Trewick SC, Koivisto P, et al. (2002) Reversal of DNA alkylation damage by two human dioxygenases. Proc Natl Acad Sci U S A 99: 16660–16665. doi: 10.1073/pnas.262589799
|
| [81] |
Trewick SC, Henshaw TF, Hausinger RP, et al. (2002) Oxidative demethylation by Escherichia coli AlkB directly reverts DNA base damage. Nature 419: 174–178. doi: 10.1038/nature00908
|
| [82] |
Tsujikawa K, Koike K, Kitae K, et al. (2007) Expression and sub-cellular localization of human ABH family molecules. J Cell Mol Med 11: 1105–1116. doi: 10.1111/j.1582-4934.2007.00094.x
|
| [83] |
Sundheim O, Talstad VA, Vagbo CB, et al. (2008) AlkB demethylases flip out in different ways. DNA Repair 7: 1916–1923. doi: 10.1016/j.dnarep.2008.07.015
|
| [84] |
Dango S, Mosammaparast N, Sowa ME, et al. (2011) DNA unwinding by ASCC3 helicase is coupled to ALKBH3-dependent DNA alkylation repair and cancer cell proliferation. Mol Cell 44: 373–384. doi: 10.1016/j.molcel.2011.08.039
|
| [85] |
Brickner JR, Soll JM, Lombardi PM, et al. (2017) A ubiquitin-dependent signalling axis specific for ALKBH-mediated DNA dealkylation repair. Nature 551: 389–393. doi: 10.1038/nature24484
|
| [86] |
Lee DH, Jin SG, Cai S, et al. (2005) Repair of methylation damage in DNA and RNA by mammalian AlkB homologues. J Biol Chem 280: 39448–39459. doi: 10.1074/jbc.M509881200
|
| [87] | Kataoka H, Yamamoto Y, Sekiguchi M (1983) A new gene (alkB) of Escherichia coli that controls sensitivity to methyl methane sulfonate. J Bacteriol 153: 1301–1307. |
| [88] |
Falnes PO, Johansen RF, Seeberg E (2002) AlkB-mediated oxidative demethylation reverses DNA damage in Escherichia coli. Nature 419: 178–182. doi: 10.1038/nature01048
|
| [89] |
Frick LE, Delaney JC, Wong C, et al. (2007) Alleviation of 1,N6-ethanoadenine genotoxicity by the Escherichia coli adaptive response protein AlkB. Proc Natl Acad Sci U S A 104: 755–760. doi: 10.1073/pnas.0607377104
|
| [90] |
Johnson RE, Yu SL, Prakash S, et al. (2007) A Role for Yeast and Human Translesion Synthesis DNA Polymerases in Promoting Replication through 3-Methyl Adenine. Mol Cell Biol 27: 7198–7205. doi: 10.1128/MCB.01079-07
|
| [91] |
Engelward BP, Allan JM, Dreslin AJ, et al. (1998) A Chemical and Genetic Approach Together Define the Biological Consequences of 3-Methyladenine Lesions in the Mammalian Genome. J Biol Chem 273: 5412–5418. doi: 10.1074/jbc.273.9.5412
|
| [92] |
Sedgwick B (2004) Repairing DNA-methylation damage. Nat Rev Mol Cell Biol 5: 148–157. doi: 10.1038/nrm1312
|
| [93] |
Shrivastav N, Li D, Essigmann JM (2010) Chemical biology of mutagenesis and DNA repair: Cellular responses to DNA alkylation. Carcinogenesis 31: 59–70. doi: 10.1093/carcin/bgp262
|
| [94] |
Furrer A, van Loon B (2014) Handling the 3-methylcytosine lesion by six human DNA polymerases members of the B-, X- and Y-families. Nucleic Acids Res 42: 553–566. doi: 10.1093/nar/gkt889
|
| [95] |
Ringvoll J, Nordstrand LM, Vagbo CB, et al. (2006) Repair deficient mice reveal mABH2 as the primary oxidative demethylase for repairing 1meA and 3meC lesions in DNA. EMBO J 25: 2189–2198. doi: 10.1038/sj.emboj.7601109
|
| [96] |
Calvo JA, Meira LB, Lee CYI, et al. (2012) DNA repair is indispensable for survival after acute inflammation. J Clin Invest 122: 2680–2689. doi: 10.1172/JCI63338
|
| [97] |
Nay SL, Lee DH, Bates SE, et al. (2012) Alkbh2 protects against lethality and mutation in primary mouse embryonic fibroblasts. DNA Repair 11: 502–510. doi: 10.1016/j.dnarep.2012.02.005
|
| [98] |
Jeggo PA, Pearl LH, Carr AM (2016) DNA repair, genome stability and cancer: A historical perspective. Nat Rev Cancer 16: 35–42. doi: 10.1038/nrc.2015.4
|
| [99] |
Park TJ, Han SU, Cho YK, et al. (2001) Methylation of O6-methylguanine-DNA methyltransferase gene is associated significantly with K-ras mutation, lymph node invasion, tumor staging, and disease free survival in patients with gastric carcinoma. Cancer 92: 2760–2768. doi: 10.1002/1097-0142(20011201)92:11<2760::AID-CNCR10123>3.0.CO;2-8
|
| [100] | Esteller M, Toyota M, Sanchez-Cespedes M, et al. (2000) Inactivation of the DNA repair gene O6-methylguanine-DNA methyltransferase by promoter hypermethylation is associated with G to A mutations in K-ras in colorectal tumorigenesis. Cancer Res 60: 2368–2371. |
| [101] | Wolf P, Hu YC, Doffek K, et al. (2001) O6-Methylguanine-DNA methyltransferase promoter hypermethylation shifts the p53 mutational spectrum in non-small cell lung cancer. Cancer Res 61: 8113–8117. |
| [102] |
Nakamura M, Watanabe T, Yonekawa Y, et al. (2001) Promoter methylation of the DNA repair gene MGMT in astrocytomas is frequently associated with G:C → A:T mutations of the TP53 tumor suppressor gene. Carcinogenesis 22: 1715–1719. doi: 10.1093/carcin/22.10.1715
|
| [103] | Esteller M, Hamilton SR, Burger PC, et al. (1999) Inactivation of the DNA repair gene O6-methylguanine-DNA methyltransferase by promoter hypermethylation is a common event in primary human neoplasia. Cancer Res 59: 793–797. |
| [104] | Sharma S, Salehi F, Scheithauer BW, et al. (2009) Role of MGMT in tumor development, progression, diagnosis, treatment and prognosis. Anticancer Res 29: 3759–3768. |
| [105] |
Soejima H, Zhao W, Mukai T (2005) Epigenetic silencing of the MGMT gene in cancer. Biochem Cell Biol 83: 429–437. doi: 10.1139/o05-140
|
| [106] |
Lee SY (2016) Temozolomide resistance in glioblastoma multiforme. Genes Dis 3: 198–210. doi: 10.1016/j.gendis.2016.04.007
|
| [107] |
Christmann M, Nagel G, Horn S, et al. (2010) MGMT activity, promoter methylation and immunohistochemistry of pretreatment and recurrent malignant gliomas: A comparative study on astrocytoma and glioblastoma. Int J Cancer 127: 2106–2118. doi: 10.1002/ijc.25229
|
| [108] |
Citron M, Schoenhaus M, Rothenberg H, et al. (1994) O6-methylguanine-DNA methyltransferase in normal and malignant tissue of the breast. Cancer Invest 12: 605–610. doi: 10.3109/07357909409023045
|
| [109] |
Musarrat J, Wilson JA, Abou-Issa H, et al. (1995) O6-alkylguanine DNA alkyltransferase activity levels in normal, benign and malignant human female breast. Biochem Biophys Res Commun 208: 688–696. doi: 10.1006/bbrc.1995.1393
|
| [110] |
Hengstler JG, Tanner B, Moller L, et al. (1999) Activity of O6-methylguanine-DNA methyltransferase in relation to p53 status and therapeutic response in ovarian cancer. Int J Cancer 84: 388–395. doi: 10.1002/(SICI)1097-0215(19990820)84:4<388::AID-IJC10>3.0.CO;2-3
|
| [111] |
Konishi N, Nakamura M, Ishida E, et al. (2005) High expression of a new marker PCA-1 in human prostate carcinoma. Clin Cancer Res 11: 5090–5097. doi: 10.1158/1078-0432.CCR-05-0195
|
| [112] |
Fedeles BI, Singh V, Delaney JC, et al. (2015) The AlkB Family of Fe(II)/alpha-Ketoglutarate-dependent Dioxygenases: Repairing Nucleic Acid Alkylation Damage and Beyond. J Biol Chem 290: 20734–20742. doi: 10.1074/jbc.R115.656462
|
| [113] |
Stefansson OA, Hermanowicz S, van der HJ, et al. (2017) CpG promoter methylation of the ALKBH3 alkylation repair gene in breast cancer. BMC Cancer 17: 469. doi: 10.1186/s12885-017-3453-8
|
| [114] |
Shimada K, Fujii T, Tsujikawa K, et al. (2012) ALKBH3 contributes to survival and angiogenesis of human urothelial carcinoma cells through NADPH oxidase and tweak/Fn14/VEGF signals. Clin Cancer Res 18: 5247–5255. doi: 10.1158/1078-0432.CCR-12-0955
|
| [115] | Krumbhaar EB, Krumbhaar HD (1919) The Blood and Bone Marrow in Yelloe Cross Gas (Mustard Gas) Poisoning: Changes produced in the Bone Marrow of Fatal Cases. J Med Res 40: 497–508. |
| [116] | Ahmad A, Nay SL, O'Connor TR, (2015) Direct Reversal Repair in Mammalian Cells, In: Chen CC, editor. Advances in DNA Repair, Rijeka: InTech, Ch. 04. |
| [117] |
Sabharwal A, Middleton MR (2006) Exploiting the role of O6-methylguanine-DNA-methyltransferase (MGMT) in cancer therapy. Curr Opin Pharmacol 6: 355–363. doi: 10.1016/j.coph.2006.03.011
|
| [118] |
Middleton MR, Margison GP (2003) Improvement of chemotherapy efficacy by inactivation of a DNA-repair pathway. Lancet Oncol 4: 37–44. doi: 10.1016/S1470-2045(03)00959-8
|
| [119] |
Dolan ME, Moschel RC, Pegg AE (1990) Depletion of mammalian O6-alkylguanine-DNA alkyltransferase activity by O6-benzylguanine provides a means to evaluate the role of this protein in protection against carcinogenic and therapeutic alkylating agents. Proc Natl Acad Sci U S A 87: 5368–5372. doi: 10.1073/pnas.87.14.5368
|
| [120] |
Pegg AE, Boosalis M, Samson L, et al. (1993) Mechanism of inactivation of human O6-alkylguanine-DNA alkyltransferase by O6-benzylguanine. Biochemistry 32: 11998–12006. doi: 10.1021/bi00096a009
|
| [121] |
Rabik CA, Njoku MC, Dolan ME (2006) Inactivation of O6-alkylguanine DNA alkyltransferase as a means to enhance chemotherapy. Cancer Treat Rev 32: 261–276. doi: 10.1016/j.ctrv.2006.03.004
|
| [122] |
Kaina B, Margison GP, Christmann M (2010) Targeting O6-methylguanine-DNA methyltransferase with specific inhibitors as a strategy in cancer therapy. Cell Mol Life Sci 67: 3663–3681. doi: 10.1007/s00018-010-0491-7
|
| [123] |
Friedman HS, Dolan ME, Moschel RC, et al. (1992) Enhancement of nitrosourea activity in medulloblastoma and glioblastoma multiforme. J Natl Cancer Inst 84: 1926–1931. doi: 10.1093/jnci/84.24.1926
|
| [124] |
Wedge SR, Porteous JK, Newlands ES (1997) Effect of single and multiple administration of an O6-benzylguanine/temozolomide combination: An evaluation in a human melanoma xenograft model. Cancer Chemother Pharmacol 40: 266–272. doi: 10.1007/s002800050657
|
| [125] | Wan Y, Wu D, Gao H, et al. (2000) Potentiation of BCNU anticancer activity by O6-benzylguanine: A study in vitro and in vivo. J Environ Pathol Toxicol Oncol 19: 69–75. |
| [126] |
Marathi UK, Dolan ME, Erickson LC (1994) Anti-neoplastic activity of sequenced administration of O6-benzylguanine, streptozotocin, and 1,3-bis(2-chloroethyl)-1-nitrosourea in vitro and in vivo. Biochem Pharmacol 48: 2127–2134. doi: 10.1016/0006-2952(94)90514-2
|
| [127] | Kreklau EL, Kurpad C, Williams DA, et al. (1999) Prolonged inhibition of O6-methylguanine DNA methyltransferase in human tumor cells by O6-benzylguanine in vitro and in vivo. J Pharmacol Exp Ther 291: 1269–1275. |
| [128] |
Ranson M, Middleton MR, Bridgewater J, et al. (2006) Lomeguatrib, a potent inhibitor of O6-alkylguanine-DNA-alkyltransferase: Phase I safety, pharmacodynamic, and pharmacokinetic trial and evaluation in combination with temozolomide in patients with advanced solid tumors. Clin Cancer Res 12: 1577–1584. doi: 10.1158/1078-0432.CCR-05-2198
|
| [129] |
Watson AJ, Middleton MR, Mcgown G, et al. (2009) O6-methylguanine-DNA methyltransferase depletion and DNA damage in patients with melanoma treated with temozolomide alone or with lomeguatrib. Br J Cancer 100: 1250–1256. doi: 10.1038/sj.bjc.6605015
|
| [130] |
Gajewski TF, Sosman J, Gerson SL, et al. (2005) Phase II trial of the O6-alkylguanine DNA alkyltransferase inhibitor O6-benzylguanine and 1,3-bis(2-chloroethyl)-1-nitrosourea in advanced melanoma. Clin Cancer Res 11: 7861–7865. doi: 10.1158/1078-0432.CCR-05-0060
|
| [131] |
Blumenthal DT, Rankin C, Stelzer KJ, et al. (2015) A Phase III study of radiation therapy (RT) and O6-benzylguanine + BCNU versus RT and BCNU alone and methylation status in newly diagnosed glioblastoma and gliosarcoma: Southwest Oncology Group (SWOG) study S0001. Int J Clin Oncol 20: 650–658. doi: 10.1007/s10147-014-0769-0
|
| [132] |
Quinn JA, Jiang SX, Reardon DA, et al. (2009) Phase II trial of temozolomide plus O6-benzylguanine in adults with recurrent, temozolomide-resistant malignant glioma. J Clin Oncol 27: 1262–1267. doi: 10.1200/JCO.2008.18.8417
|
| [133] |
Friedman HS, Pluda J, Quinn JA, et al. (2000) Phase I trial of carmustine plus O6-benzylguanine for patients with recurrent or progressive malignant glioma. J Clin Oncol 18: 3522–3528. doi: 10.1200/JCO.2000.18.20.3522
|
| [134] |
Khan O, Middleton MR (2007) The therapeutic potential of O6-alkylguanine DNA alkyltransferase inhibitors. Expert Opin Invest Drugs 16: 1573–1584. doi: 10.1517/13543784.16.10.1573
|
| [135] |
Tomaszowski KH, Hellmann N, Ponath V, et al. (2017) Uptake of glucose-conjugated MGMT inhibitors in cancer cells: Role of flippases and type IV P-type ATPases. Sci Rep 7: 13925. doi: 10.1038/s41598-017-14129-x
|
| [136] |
Javanmard S, Loktionova NA, Fang Q, et al. (2007) Inactivation of O6-alkylguanine-DNA alkyltransferase by folate esters of O6-benzyl-2'-deoxyguanosine and of O6-[4-(hydroxymethyl)benzyl]guanine. J Med Chem 50: 5193–5201. doi: 10.1021/jm0705859
|
| [137] |
Goder A, Nagel G, Kraus A, et al. (2015) Lipoic acid inhibits the DNA repair protein O 6-methylguanine-DNA methyltransferase (MGMT) and triggers its depletion in colorectal cancer cells with concomitant autophagy induction. Carcinogenesis 36: 817–831. doi: 10.1093/carcin/bgv070
|
| [138] |
Wang K, Chen D, Qian Z, et al. (2017) Hedgehog/Gli1 signaling pathway regulates MGMT expression and chemoresistance to temozolomide in human glioblastoma. Cancer Cell Int 17: 117. doi: 10.1186/s12935-017-0491-x
|
| [139] |
Wickstrom M, Dyberg C, Milosevic J, et al. (2015) Wnt/beta-catenin pathway regulates MGMT gene expression in cancer and inhibition of Wnt signalling prevents chemoresistance. Nat Commun 6: 8904. doi: 10.1038/ncomms9904
|
| [140] |
Esteller M, Herman JG (2004) Generating mutations but providing chemosensitivity: The role of O6-methylguanine DNA methyltransferase in human cancer. Oncogene 23: 1–8. doi: 10.1038/sj.onc.1207316
|
| [141] | Nakada M, Furuta T, Hayashi Y, et al. (2012) The strategy for enhancing temozolomide against malignant glioma. Front Oncol 2: 98. |
| [142] |
Moen EL, Stark AL, Zhang W, et al. (2014) The role of gene body cytosine modifications in MGMT expression and sensitivity to temozolomide. Mol Cancer Ther 13: 1334–1344. doi: 10.1158/1535-7163.MCT-13-0924
|
| [143] |
Krumm A, Barckhausen C, Kucuk P, et al. (2016) Enhanced histone deacetylase activity in malignant melanoma provokes RAD51 and FANCD2-triggered drug resistance. Cancer Res 76: 3067–3077. doi: 10.1158/0008-5472.CAN-15-2680
|
| [144] | Ryu CH, Yoon WS, Park KY, et al. (2012) Valproic acid downregulates the expression of MGMT and sensitizes temozolomide-resistant glioma cells. J Biomed Biotechnol 2012: 987495. |
| [145] | Li Z, Xia Y, Bu X, et al. (2018) Effects of valproic acid on the susceptibility of human glioma stem cells for TMZ and ACNU. Oncol Lett 15: 9877–9883. |
| [146] |
Eckert M, Klumpp L, Huber SM (2017) Cellular Effects of the Antiepileptic Drug Valproic Acid in Glioblastoma. Cell Physiol Biochem 44: 1591–1605. doi: 10.1159/000485753
|
| [147] |
Johannessen TC, Prestegarden L, Grudic A, et al. (2013) The DNA repair protein ALKBH2 mediates temozolomide resistance in human glioblastoma cells. Neuro Oncol 15: 269–278. doi: 10.1093/neuonc/nos301
|
| [148] |
Komander D, Rape M (2012) The ubiquitin code. Annu Rev Biochem 81: 203–229. doi: 10.1146/annurev-biochem-060310-170328
|
| [149] |
Zhao Y, Majid MC, Soll JM, et al. (2015) Noncanonical regulation of alkylation damage resistance by the OTUD4 deubiquitinase. EMBO J 34: 1687–1703. doi: 10.15252/embj.201490497
|
| [150] |
Chauhan D, Tian Z, Nicholson B, et al. (2012) A small molecule inhibitor of ubiquitin-specific protease-7 induces apoptosis in multiple myeloma cells and overcomes bortezomib resistance. Cancer Cell 22: 345–358. doi: 10.1016/j.ccr.2012.08.007
|
| [151] |
Kapuria V, Peterson LF, Fang D, et al. (2010) Deubiquitinase inhibition by small-molecule WP1130 triggers aggresome formation and tumor cell apoptosis. Cancer Res 70: 9265–9276. doi: 10.1158/0008-5472.CAN-10-1530
|
| [152] |
Wang P, Wu J, Shenghong M, et al. (2015) Oncometabolite D-2-hydroxyglutarate inhibits ALKBH DNA repair enzymes and sensitizes IDH-mutant cells to alkylating agents. Cell Rep 13: 2353–2361. doi: 10.1016/j.celrep.2015.11.029
|
| [153] |
Tran TQ, Ishak Gabra MB, Lowman XH, et al. (2017) Glutamine deficiency induces DNA alkylation damage and sensitizes cancer cells to alkylating agents through inhibition of ALKBH enzymes. PLOS Biol 15: e2002810. doi: 10.1371/journal.pbio.2002810
|
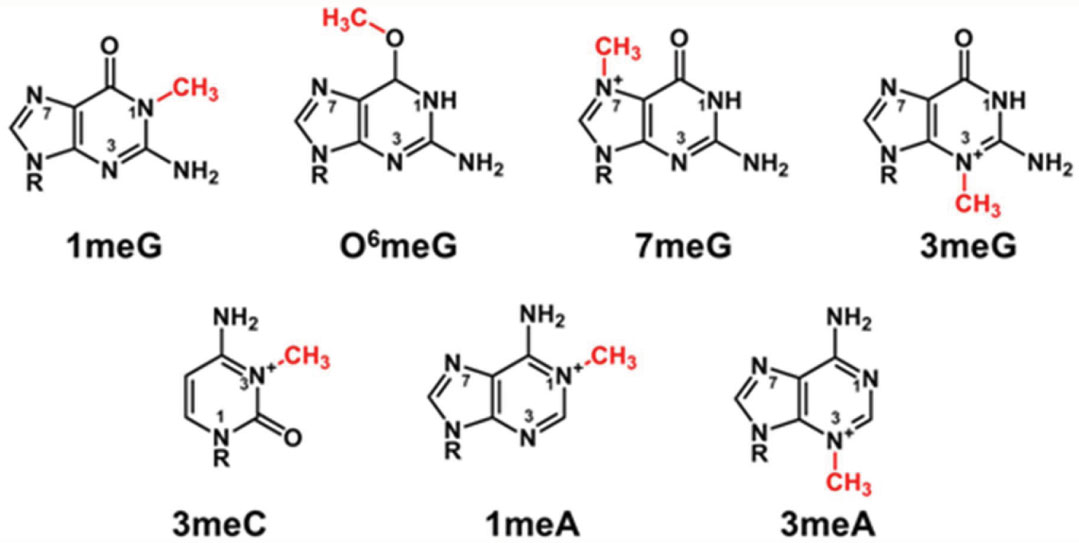
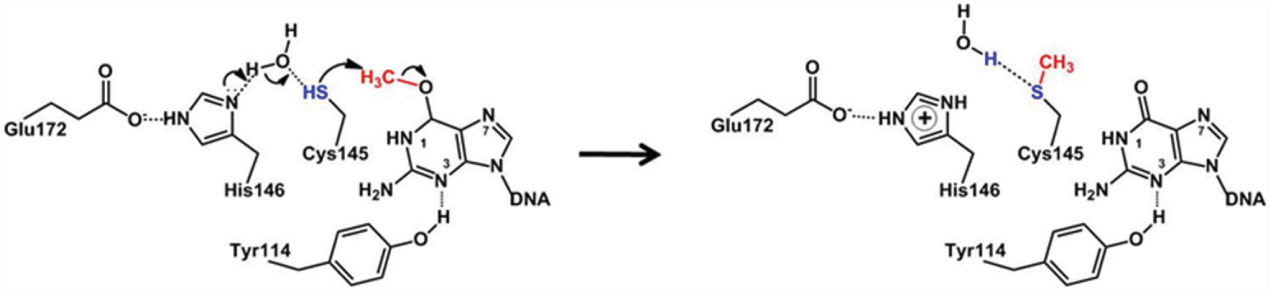
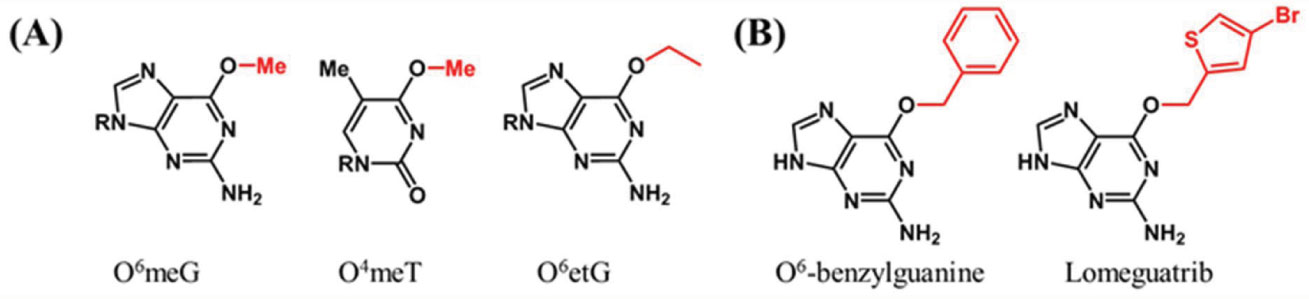
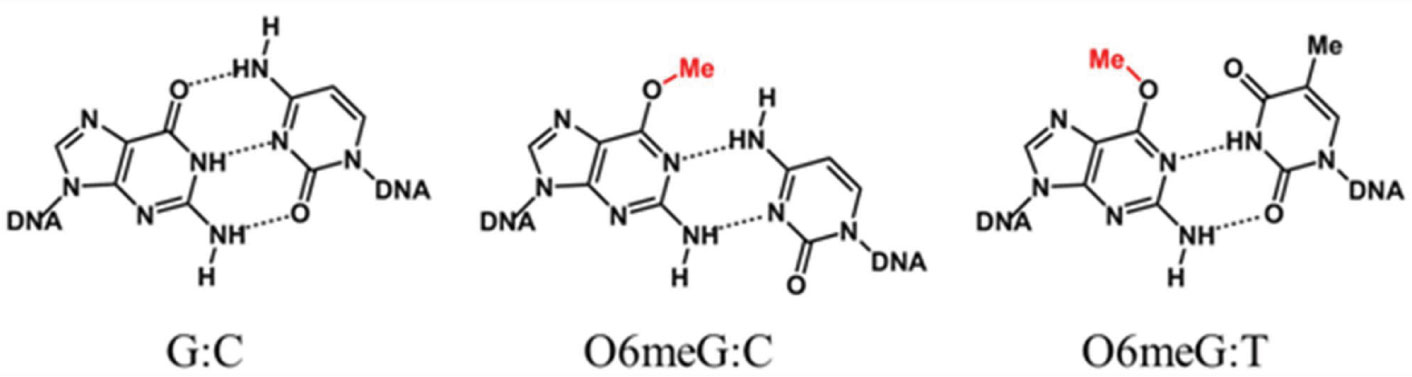
Figures(7)

Roberto Gutierrez, Yesenia Thompson, Timothy R. O’Connor. DNA direct repair pathways in cancer[J]. AIMS Medical Science, 2018, 5(3): 284-302. doi: 10.3934/medsci.2018.3.284







 DownLoad:
DownLoad: