This paper discusses the robustness of neutral-type Cohen-Grossberg neural networks with time delays and stochastic disturbances. And the problem is whether the Cohen-Grossberg neural networks, which originally maintain exponential stability, still achieves exponential stability when subjected to three simultaneous disturbances, namely, time delays, stochastic perturbations, and neutral terms. First, the width of the time delays, the strength of the stochastic disturbances, and the neutral term preset parameter size are derived through the Bellman-Gronwall Lemma, the Itô formula, and the properties of integrals. Next, the values of the three perturbation factors of time delay, stochastic disturbance, and neutral term are obtained by solving a multivariate privacy transcendental equation, which allows the Cohen-Grossberg neural networks to remain exponentially stable after being disturbed. Finally, the numerical example is provided to validate the results of this brief.
Citation: Yijia Zhang, Tao Xie, Yunlong Ma. Robustness analysis of exponential stability of Cohen-Grossberg neural network with neutral terms[J]. AIMS Mathematics, 2025, 10(3): 4938-4954. doi: 10.3934/math.2025226
This paper discusses the robustness of neutral-type Cohen-Grossberg neural networks with time delays and stochastic disturbances. And the problem is whether the Cohen-Grossberg neural networks, which originally maintain exponential stability, still achieves exponential stability when subjected to three simultaneous disturbances, namely, time delays, stochastic perturbations, and neutral terms. First, the width of the time delays, the strength of the stochastic disturbances, and the neutral term preset parameter size are derived through the Bellman-Gronwall Lemma, the Itô formula, and the properties of integrals. Next, the values of the three perturbation factors of time delay, stochastic disturbance, and neutral term are obtained by solving a multivariate privacy transcendental equation, which allows the Cohen-Grossberg neural networks to remain exponentially stable after being disturbed. Finally, the numerical example is provided to validate the results of this brief.

| [1] |
M. A. Cohen, S. Grossberg, Absolute stability of global pattern formation and parallel memory storage by competitive neural networks, IEEE T. Syst. Man Cy., 1983,815–826. http://dx.doi.org/10.1109/tsmc.1983.6313075 doi: 10.1109/tsmc.1983.6313075

|
| [2] |
C. J. Cheng, T. L. Liao, J. J. Yan, C. C. Hwang, Globally asymptotic stability of a class of neutral-type neural networks with delays, IEEE T. Syst. Man Cy. B, 36 (2006), 1191–1195. http://dx.doi.org/10.1109/TSMCB.2006.874677 doi: 10.1109/TSMCB.2006.874677
|
| [3] |
P. Kowsalya, S. S. Mohanrasu, A. Kashkynbayev, P. Gokul, R. Rakkiyappan, Fixed-time synchronization of inertial Cohen-Grossberg neural networks with state dependent delayed impulse control and its application to multi-image encryption, Chaos Soliton. Fract., 181 (2024), 114693. http://dx.doi.org/10.1016/j.chaos.2024.114693 doi: 10.1016/j.chaos.2024.114693
|
| [4] |
J. Li, Z. Peng, Multi-source image fusion algorithm based on cellular neural networks with genetic algorithm, Optik, 126 (2015), 5230–5236. http://dx.doi.org/10.1016/j.ijleo.2015.09.187 doi: 10.1016/j.ijleo.2015.09.187
|
| [5] |
X. Hu, G. Feng, S. Duan, L. Liu, A memristive multilayer cellular neural network with applications to image processing, IEEE T. Neur. Net. Lear., 28 (2016), 1889–1901. http://dx.doi.org/10.1109/TNNLS.2016.2552640 doi: 10.1109/TNNLS.2016.2552640
|
| [6] |
X. Yang, Z. Cheng, X. Li, T. Ma, Exponential synchronization of coupled neutral-type neural networks with mixed delays via quantized output control, J. Franklin I., 356 (2019), 8138–8153. http://dx.doi.org/10.1016/j.jfranklin.2019.07.006 doi: 10.1016/j.jfranklin.2019.07.006
|
| [7] |
R. Samli, S. Senan, E. Yucel, Z. Orman, Some generalized global stability criteria for delayed Cohen–Grossberg neural networks of neutral-type, Chaos Soliton. Fract., 116 (2019), 198–207. http://dx.doi.org/10.1016/j.neunet.2019.04.023 doi: 10.1016/j.neunet.2019.04.023
|
| [8] |
Z. Zhang, X. Zhang, T. Yu, Global exponential stability of neutral-type Cohen–Grossberg neural networks with multiple time-varying neutral and discrete delays, Neurocomputing, 490 (2022), 124–131. http://dx.doi.org/10.1016/j.neucom.2022.03.068 doi: 10.1016/j.neucom.2022.03.068
|
| [9] |
F. Kong, Q. Zhu, Antiperiodic dynamical behaviors of discontinuous neutral-type Cohen-Grossberg neural networks with mixed time delays, Comput. Intel., 36 (2020), 698–719. http://dx.doi.org/10.1111/coin.12262 doi: 10.1111/coin.12262
|
| [10] |
Y. Shen, J. Wang, Robustness analysis of global exponential stability of non-linear systems with time delays and neutral terms, IET Control Theory A., 7 (2013), 1227–1232. http://dx.doi.org/10.1049/iet-cta.2012.0781 doi: 10.1049/iet-cta.2012.0781
|
| [11] |
O. Faydasicok, S. Arik, The combined Lyapunov functionals method for stability analysis of neutral Cohen-Grossberg neural networks with multiple delays, Neural Networks, 180 (2024), 106641. http://dx.doi.org/10.1016/j.neunet.2024.106641 doi: 10.1016/j.neunet.2024.106641
|
| [12] |
R. Li, J. Cao, A. Alsaedi, F. Alsaadi, Exponential and fixed-time synchronization of Cohen–Grossberg neural networks with time-varying delays and reaction-diffusion terms, Appl. Math. Comput., 313 (2017), 37–51. http://dx.doi.org/10.1016/j.amc.2017.05.073 doi: 10.1016/j.amc.2017.05.073
|
| [13] |
Y. Wan, J. Cao, G. Wen, W. Yu, Robust fixed-time synchronization of delayed Cohen–Grossberg neural networks, Neural Networks, 73 (2016), 86–94. http://dx.doi.org/10.1016/j.neunet.2015.10.009 doi: 10.1016/j.neunet.2015.10.009
|
| [14] |
N. Ozcan, New conditions for global stability of neutral-type delayed Cohen–Grossberg neural networks, Neural Networks, 106 (2018), 1–7. http://dx.doi.org/10.1016/j.neunet.2018.06.009 doi: 10.1016/j.neunet.2018.06.009
|
| [15] |
H. Zhang, Inequalities and stability of stochastic fuzzy delayed Cohen–Grossberg neural networks, IEEE Access, 2023. http://dx.doi.org/10.1109/ACCESS.2023.3300581 doi: 10.1109/ACCESS.2023.3300581
|
| [16] |
X. Z. Liu, K. N. Wu, X. Ding, W. Zhang, Boundary stabilization of stochastic delayed Cohen–Grossberg neural networks with diffusion terms, IEEE T. Neur. Net. Lear., 33 (2021), 3227–3237. http://dx.doi.org/10.1109/TNNLS.2021.3051363 doi: 10.1109/TNNLS.2021.3051363
|
| [17] | M. A. Jamal, S. Das, S. Mukhopadhyay, Fixed-time synchronization of delayed inertial Cohen–Grossberg neural networks with desynchronizing impulses, Commun. Nonlinear Sci., 130 (2024), 107772. http://dx.doi.org/0.1016/j.cnsns.2023.107772 |
| [18] |
J. Cheng, W. Liu, Stability analysis of anti-periodic solutions for Cohen–Grossberg neural networks with inertial term and time delays, Mathematics, 12 (2024), 198. http://dx.doi.org/10.3390/math12020198 doi: 10.3390/math12020198
|
| [19] |
M. A. Jamal, R. Kumar, S. Mukhopadhyay, O. M. Kwon, Fixed-time stability of Cohen-Grossberg BAM neural networks with impulsive perturbations, Neurocomputing, 550 (2023), 126501. http://dx.doi.org/10.1016/j.neucom.2023.126501 doi: 10.1016/j.neucom.2023.126501
|
| [20] |
X. Wang, J. Lan, X. Yang, X. Zhang, Global robust exponential synchronization of neutral-type interval Cohen-Grossberg neural networks with mixed time delays, Inform. Sciences, 676 (2024), 120806. http://dx.doi.org/10.1016/j.ins.2024.120806 doi: 10.1016/j.ins.2024.120806
|
| [21] |
M. Shen, Z. Wang, S. Zhu, X. Zhao, G. Zong, Q. G. Wang, Neural network adaptive iterative learning control for strict-feedback unknown delay systems against input saturation, IEEE T. Neur. Net. Lear., 2024, 1–10. http://dx.doi.org/10.1109/TNNLS.2024.3452721 doi: 10.1109/TNNLS.2024.3452721
|
| [22] |
Q. Meng, Q. Ma, Y. Shi, Adaptive fixed-time stabilization for a class of uncertain nonlinear systems, IEEE T. Automatic Contr., 68 (2023), 6929–6936. http://dx.doi.org/10.1109/TAC.2023.3244151 doi: 10.1109/TAC.2023.3244151
|
| [23] |
L. Xing, C. Wen, Dynamic event-triggered adaptive control for a class of uncertain nonlinear systems, Automatica, 158 (2023), 111286. http://dx.doi.org/10.1016/j.automatica.2023.111286 doi: 10.1016/j.automatica.2023.111286
|
| [24] |
M. Shen, C. Wang, Q. G. Wang, Y. Sun, G. Zong, Synchronization of fractional reaction-diffusion complex networks with unknown couplings, IEEE T. Netw. Sci. Eng., 11 (2024), 4503–4512. http://dx.doi.org/10.1109/TNSE.2024.3432997 doi: 10.1109/TNSE.2024.3432997
|
| [25] |
H. Liu, J. Cheng, J. Cao, I. Katib, Preassigned-time synchronization for complex-valued memristive neural networks with reaction–diffusion terms and Markov parameters, Neural Networks, 169 (2024), 520–531. http://dx.doi.org/10.1016/j.neunet.2023.11.011 doi: 10.1016/j.neunet.2023.11.011
|
| [26] |
W. Fang, T. Xie, B. Li, Robustness analysis of fuzzy cellular neural network with deviating argument and stochastic disturbances, IEEE Access, 11 (2023), 3717–3728. http://dx.doi.org/10.1109/ACCESS.2023.3233946 doi: 10.1109/ACCESS.2023.3233946
|
| [27] |
S. Majhi, B. Rakshit, A. Sharma, J. Kurths, D. Ghosh, Dynamical robustness of network of oscillators, Phys. Rep., 1082 (2024), 1–46. http://dx.doi.org/10.1016/j.physrep.2024.06.003 doi: 10.1016/j.physrep.2024.06.003
|
| [28] |
O. Artime, M. Grassia, M. Domenico, J. P. Gleeson, H. A. Makse, G. Mangioni, Robustness and resilience of complex networks, Nat. Rev. Phys., 6 (2024), 114–131. http://dx.doi.org/10.1038/s42254-023-00676-y doi: 10.1038/s42254-023-00676-y
|
| [29] |
W. Si, S. Gao, F. Yan, N. Zhao, H. Zhang, H. Dong, Linkage-constraint criteria for robust exponential stability of nonlinear BAM system with derivative contraction coefficients and piecewise constant arguments, Inform. Sciences, 612 (2022), 926–941. http://dx.doi.org/10.1016/j.ins.2022.08.078 doi: 10.1016/j.ins.2022.08.078
|
| [30] |
Y. Zhang, D. W. Gu, S. Xu, Global exponential adaptive synchronization of complex dynamical networks with neutral-type neural network nodes and stochastic disturbances, IEEE T. Circuits I, 60 (2013), 2709–2718. http://dx.doi.org/10.1109/TCSI.2013.2249151 doi: 10.1109/TCSI.2013.2249151
|
| [31] |
O. Faydasicok, S. Arik, An analysis of stability of multiple delayed Cohen-Grossberg neural networks of neutral type, 2022 7th International Conference on Mathematics and Computers in Sciences and Industry (MCSI), 2022, 55–60. http://dx.doi.org/10.1109/MCSI55933.2022.00016 doi: 10.1109/MCSI55933.2022.00016
|
| [32] |
L. Wan, Q. Zhou, Stability analysis of neutral-type Cohen-Grossberg neural networks with multiple time-varying delays, IEEE Access, 8 (2020), 27618–27623. http://dx.doi.org/10.1109/ACCESS.2020.2971839 doi: 10.1109/ACCESS.2020.2971839
|
| [33] |
L. Wan, Q. Zhou, Exponential stability of neutral-type Cohen-Grossberg neural networks with multiple time-varying delays, IEEE Access, 9 (2021), 48914–48922. http://dx.doi.org/10.1109/ACCESS.2021.3068191 doi: 10.1109/ACCESS.2021.3068191
|
| [34] |
W. Yao, C. Wang, Y. Sun, C. Zhou, H. Lin, Exponential multistability of memristive Cohen-Grossberg neural networks with stochastic parameter perturbations, Appl. Math. Comput., 386 (2020), 125483. http://dx.doi.org/10.1016/j.amc.2020.125483 doi: 10.1016/j.amc.2020.125483
|
| [35] |
Q. Zhu, J. Cao, Robust exponential stability of Markovian jump impulsive stochastic Cohen-Grossberg neural networks with mixed time delays, IEEE T. Neural Networks, 21 (2010), 1314–1325. http://dx.doi.org/10.1109/TNN.2010.2054108 doi: 10.1109/TNN.2010.2054108
|
| [36] |
Y. Shen, J. Wang, Robustness of global exponential stability of nonlinear systems with random disturbances and time delays, IEEE T. Syst. Man Cy.-S., 46 (2015), 1157–1166. http://dx.doi.org/10.1109/TSMC.2015.2497208 doi: 10.1109/TSMC.2015.2497208
|
| [37] |
N. Ozcan, Stability analysis of Cohen–Grossberg neural networks of neutral-type: Multiple delays case, Neural Networks, 113 (2019), 20–27. http://dx.doi.org/10.1016/j.neunet.2019.01.017 doi: 10.1016/j.neunet.2019.01.017
|
| [38] | X. Mao, Stochastic differential equations and applications, 2 Eds., UK: Woodhead Publishing, 2008. |
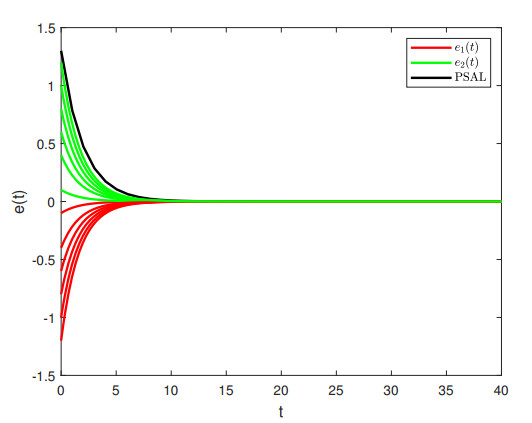



Figures(4) / Tables(1)

Yijia Zhang, Tao Xie, Yunlong Ma. Robustness analysis of exponential stability of Cohen-Grossberg neural network with neutral terms[J]. AIMS Mathematics, 2025, 10(3): 4938-4954. doi: 10.3934/math.2025226







 DownLoad:
DownLoad: