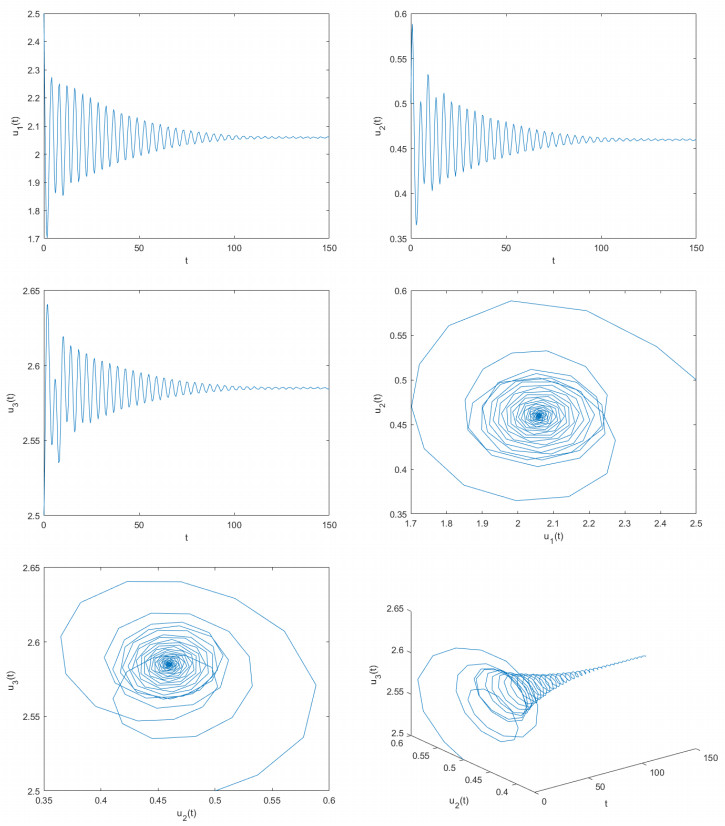
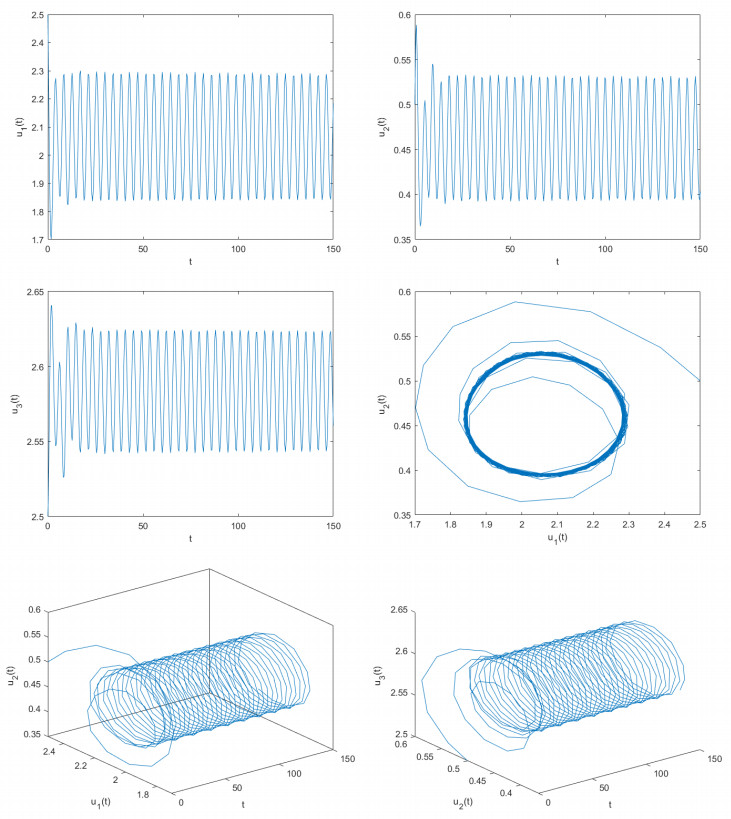
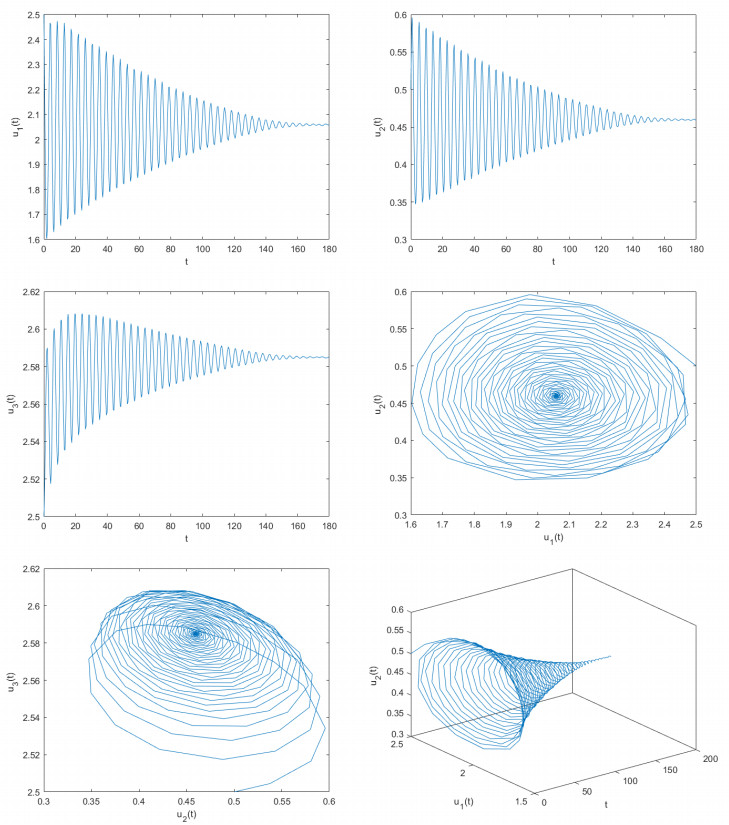
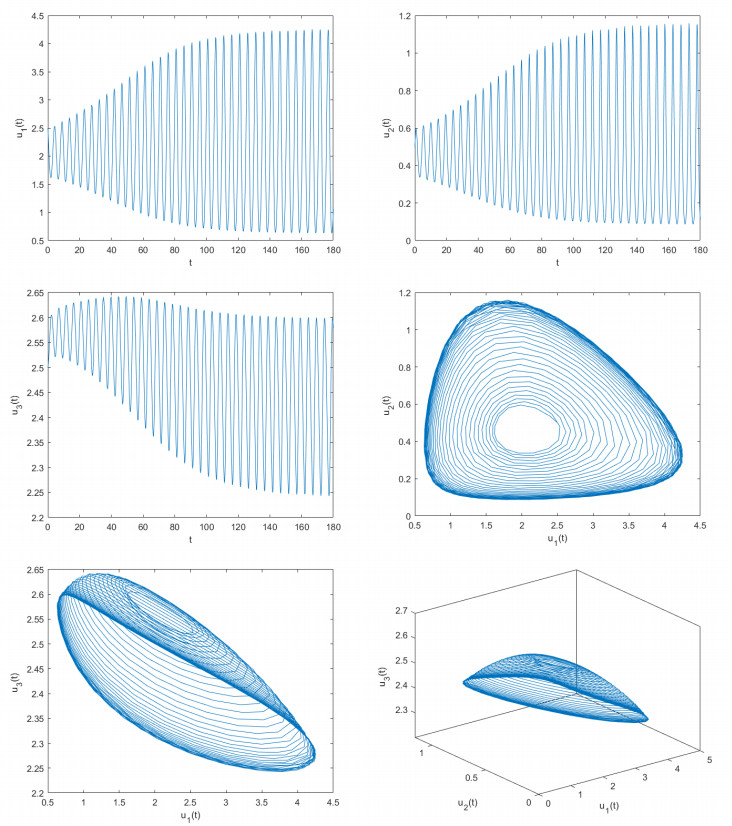
Delayed dynamical models demonstrate significant application value in depicting interactions and internal dynamics among different biological populations. Therefore, they have garnered significant interest from numerous scholars in both biology and mathematics. Based on previous studies, this article construct a novel delayed predator-prey model. By utilizing fixed point theory, inequality methods, and appropriate functions, this article examined the desirable properties of the solutions of the constructed delayed predator-prey system, including existence and uniqueness, boundedness, and non-negativity. This paper determines the parameter conditions for system stability and the occurrence of bifurcations by employing bifurcation theory and the stability theory of delayed differential equations. Using two control strategies, namely the mixed controller and the extended delay feedback controller, this paper effectively adjusts the stability domain of the delayed predator-prey systems and controls the time of bifurcation onset. The research explores how delays affect the stabilization of system and the adjustment of bifurcation. This paper provides computer simulation photos supporting the main obtained findings. The outcomes of this paper are groundbreaking and can provide critical guidance for the control and regulation of predator and prey population densities.
Citation: Jinting Lin, Changjin Xu, Yiya Xu, Yingyan Zhao, Yicheng Pang, Zixin Liu, Jianwei Shen. Bifurcation and controller design in a 3D delayed predator-prey model[J]. AIMS Mathematics, 2024, 9(12): 33891-33929. doi: 10.3934/math.20241617
Delayed dynamical models demonstrate significant application value in depicting interactions and internal dynamics among different biological populations. Therefore, they have garnered significant interest from numerous scholars in both biology and mathematics. Based on previous studies, this article construct a novel delayed predator-prey model. By utilizing fixed point theory, inequality methods, and appropriate functions, this article examined the desirable properties of the solutions of the constructed delayed predator-prey system, including existence and uniqueness, boundedness, and non-negativity. This paper determines the parameter conditions for system stability and the occurrence of bifurcations by employing bifurcation theory and the stability theory of delayed differential equations. Using two control strategies, namely the mixed controller and the extended delay feedback controller, this paper effectively adjusts the stability domain of the delayed predator-prey systems and controls the time of bifurcation onset. The research explores how delays affect the stabilization of system and the adjustment of bifurcation. This paper provides computer simulation photos supporting the main obtained findings. The outcomes of this paper are groundbreaking and can provide critical guidance for the control and regulation of predator and prey population densities.

| [1] |
Y. Enatsu, J. Roy, M. Banerjee, Hunting cooperation in a prey-predator model with maturation delay, J. Biol. Dyn., 18 (2024), 2332279. http://dx.doi.org/10.1080/17513758.2024.2332279 doi: 10.1080/17513758.2024.2332279

|
| [2] |
P. Majumdar, B. Mondal, S. Debnath, S. Sarkar, U. Ghosh, Effect of fear and delay on a prey-predator model with predator harvesting, Appl. Math. Comput., 41 (2022), 1–36. http://dx.doi.org/10.1007/s40314-022-02066-z doi: 10.1007/s40314-022-02066-z
|
| [3] |
H. Gao, X. Li, Hopf bifurcation of predator-prey symbiotic model with time delay, J. Jilin. Univ., 61 (2023), 1339–1350. http://dx.doi.org/10.13413/j.cnki.jdxblxb.2023043 doi: 10.13413/j.cnki.jdxblxb.2023043
|
| [4] |
Y. W. Yang, D. Y. Wu, C. S. Shen, J. Gao, F. P. Lu, Impacts of fear effect and nonlocal competition on a diffusive prey-predator model with delay, J. Appl. Anal. Comput., 69 (2023), 2155–2176. http://dx.doi.org/10.1007/s12190-022-01827-7 doi: 10.1007/s12190-022-01827-7
|
| [5] |
X. Y. Zhou, Stability and Hopf bifurcation analysis of a stage-structured predator-prey model with delay, Axioms, 11 (2022), 575. http://dx.doi.org/10.3390/axioms11100575 doi: 10.3390/axioms11100575
|
| [6] |
G. Baier, S. Sahle, Hyperchaos and chaotic hierarchy in low-dimensional chemical systems, JCP., 100 (1994), 8907–8911. http://dx.doi.org/10.1063/1.466694 doi: 10.1063/1.466694
|
| [7] |
B. Lakpathendup, B. Samir, K. Tapankumar, Dynamical analysis of delayed predator-prey models and explicit impacts of harvesting, J. Nonlinear Sci. Appl., 13 (2024), 373–403. http://dx.doi.org/10.5890/JAND.2024.06.013 doi: 10.5890/JAND.2024.06.013
|
| [8] |
S. Pal, P. K. Tiwari, A. K. Misra, H. Wang, Fear effect in a three-species food chain model with generalist predator, Math. Bio. Eng., 21 (2024), 1–13. http://dx.doi.org/10.3934/mbe.2024001 doi: 10.3934/mbe.2024001
|
| [9] |
C. J. Xu, X. H. Cui, P. L. Li, J. L. Yan, L. Y. Yao, Exploration on dynamics in a discrete predator-prey competitive model involving time delays and feedback controls, J. Biol. Dyn., 17 (2023), 2220349. http://dx.doi.org/10.1080/17512758.2023.2220349 doi: 10.1080/17512758.2023.2220349
|
| [10] |
P. L. Li, R. Gao, C. J. Xu, J. W. Shen, S. Ahmad, Y. Li, Exploring the impact of delay on Hopf bifurcation of a type of BAM neural network models concerning three nonidentical delays, Neural Process. Lett., 55 (2023), 11595–11635. http://dx.doi.org/10.1007/s11063-023-11392-0 doi: 10.1007/s11063-023-11392-0
|
| [11] |
Q. Y. Cui, C. J. Xu, W. Ou, Y. C. Pang, Z. X. Liu, P. L. Li, et al., Bifurcation behavior and hybrid controller design of a 2D Lotka-Volterra commensal symbiosis system accompanying delay, AIMS Math., 11 (2023), 4808. http://dx.doi.org/10.3390/math11234808 doi: 10.3390/math11234808
|
| [12] |
L. Z. Fei, X. W. Chen, B. S. Han, Bifurcation analysis and hybrid control of a discrete-time predator-prey model, J. Differ. Equ. Appl., 27 (2021), 102–117. http://dx.doi.org/10.1080/10236198.2021.1876038 doi: 10.1080/10236198.2021.1876038
|
| [13] |
K. Chakraborty, M. Chakraborty, T. K. Kar, Bifurcation and control of a bioeconomic model of a prey-predator system with a time delay, Nonlinear. Anal-Hybri., 5 (2021), 613–625. http://dx.doi.org/10.1016/j.nahs.2011.05.004 doi: 10.1016/j.nahs.2011.05.004
|
| [14] |
Z. R. Liu, K. W. Chung, Hybrid control of bifurcation in continuous nonlinear dynamical systems, Int. J. Bifurcat. Chaos., 15 (2005), 3895–3903. http://dx.doi.org/10.1142/S0218127405014374 doi: 10.1142/S0218127405014374
|
| [15] | Y. Y. Zhao, C. J. Xu, Y. Y. Xu, J. T. Lin, Y. C. Pang, Z. X. Liu, et al., Mathematical exploration on control of bifurcation for a 3D predatorprey model with delay, AIMS Math., 9 (20024), 29883–29915. http://dx.doi.org/10.3934/math.20241445 |
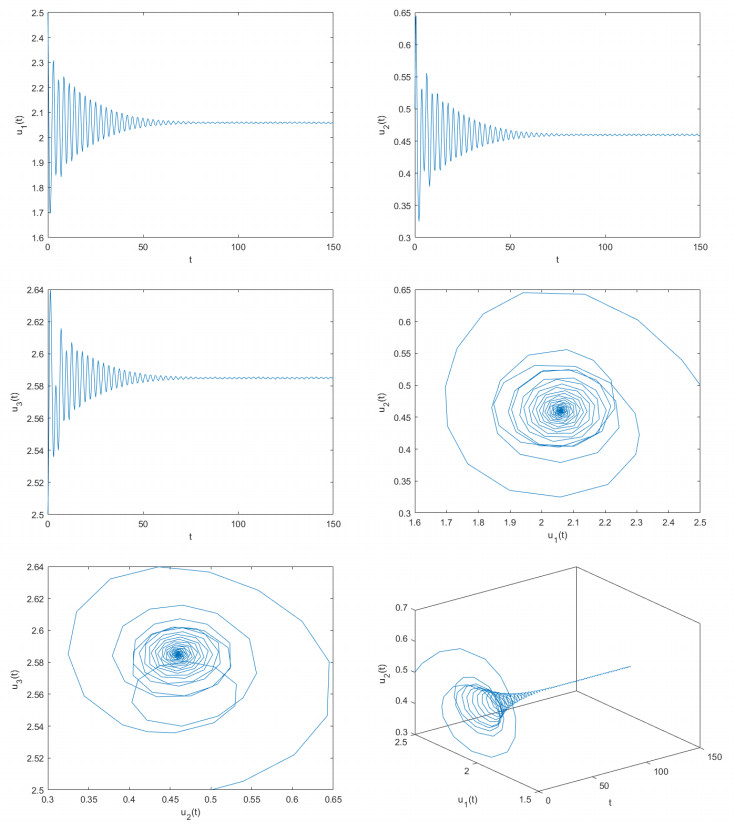
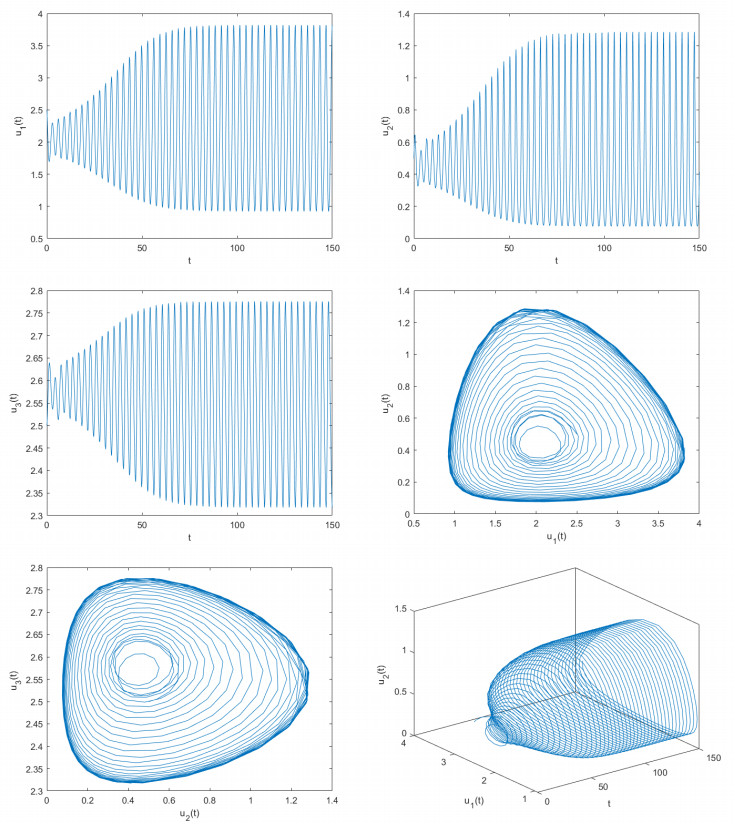
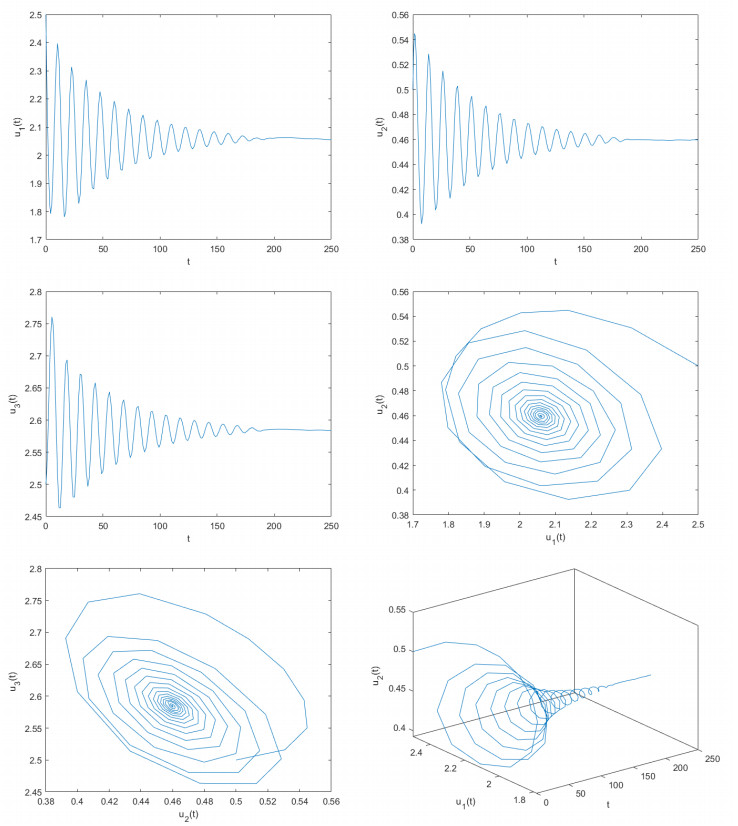
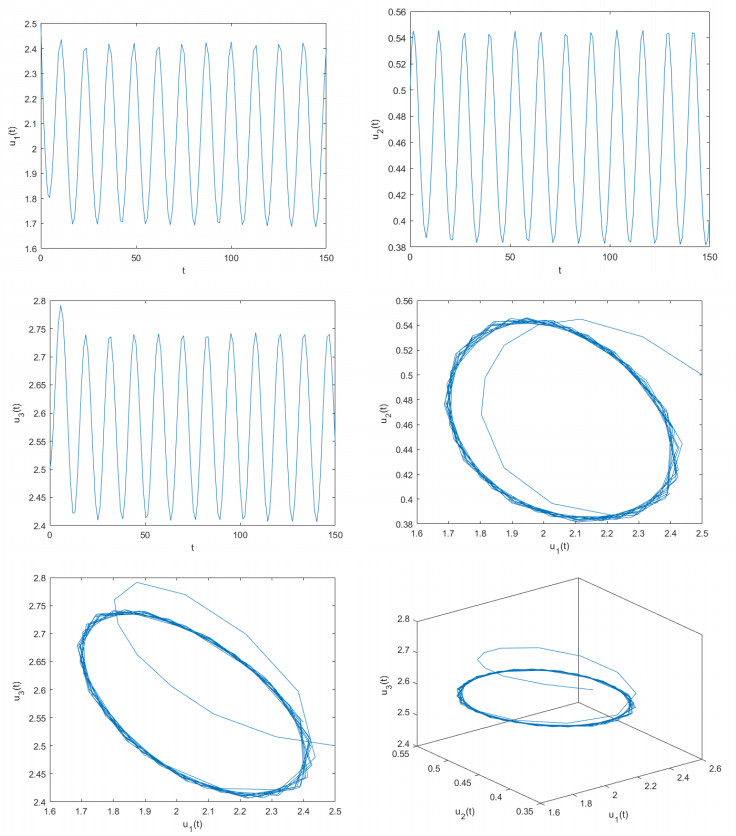
Figures(8)

Jinting Lin, Changjin Xu, Yiya Xu, Yingyan Zhao, Yicheng Pang, Zixin Liu, Jianwei Shen. Bifurcation and controller design in a 3D delayed predator-prey model[J]. AIMS Mathematics, 2024, 9(12): 33891-33929. doi: 10.3934/math.20241617







 DownLoad:
DownLoad: