Citation: Zareen Gul, Muhammad Younas Khan Barozai, Muhammad Din. In-silico based identification and functional analyses of miRNAs and their targets in Cowpea (Vigna unguiculata L.)[J]. AIMS Genetics, 2017, 4(2): 138-165. doi: 10.3934/genet.2017.2.138

| [1] |
Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281-297. doi: 10.1016/S0092-8674(04)00045-5

|
| [2] |
Kim YJ, Zheng B, Yu Y, et al. (2011) The role of mediator in small and long noncoding RNA production in Arabidopsis thaliana. EMBO J 30: 814-822. doi: 10.1038/emboj.2011.3
|
| [3] |
Zhang BH, Pan XP, Wang QL, et al. (2005) Identification and characterization of new plant microRNAs using EST analysis. Cell Res 15: 336-360. doi: 10.1038/sj.cr.7290302
|
| [4] |
Hammond SC, Bernstein E, Beach D, et al. (2000) An RNA-directed nuclease mediates posttranscriptional gene silencing in Drosophila cells. Nature 404: 293-296. doi: 10.1038/35005107
|
| [5] |
Kidner CA, Martienssen RA (2005) The developmental role of microRNA in plants. Curr Opin Plant Biol 8: 38-44. doi: 10.1016/j.pbi.2004.11.008
|
| [6] |
Baloch IA, Barozai MYK, Din M (2013) MicroRNAs: the mega regulators in eukaryotic genomes. Pure Appl Biol 2: 83-88. doi: 10.19045/bspab.2013.23002
|
| [7] |
Bai M, Yang GS, Chen WT, et al. (2012) Genome-wide identification of Dicer-like, Argonaute and RNA dependent RNA polymerase gene families and their expression analyses in response to viral infection and abiotic stresses in Solanum lycopersicum. Gene 501: 52-62. doi: 10.1016/j.gene.2012.02.009
|
| [8] | Barozai MYK (2012) Insilico identification of microRNAs and their targets in fiber and oil producing plant Flax (Linum usitatissimum L.). Pak J Bot 44: 1357-1362. |
| [9] |
Gao P, Bai X, Yang L, et al. (2011) Osa-MIR393: a salinity- and alkaline stress-related microRNA gene. Mol Biol Rep 38: 237-242. doi: 10.1007/s11033-010-0100-8
|
| [10] |
Shui XR, Chen ZW, Li JX (2013) MicroRNA prediction and its function in regulating drought-related genes in cowpea. Plant Sci 210: 25-35. doi: 10.1016/j.plantsci.2013.05.002
|
| [11] |
Xie FL, Huang SQ, Guo K, et al. (2007) Computational identification of novel microRNAs and targets in Brassica napus. FEBS Lett 581: 1464-1474. doi: 10.1016/j.febslet.2007.02.074
|
| [12] |
Zhang BH, Pan XP, Stellwag EJ (2008) Identification of soybean microRNAs and their targets. Planta 229: 161-182. doi: 10.1007/s00425-008-0818-x
|
| [13] |
Barozai MYK, Irfan M, Yousaf R, et al. (2008) Identification of micro-RNAs in cotton. Plant Physiol Biochem 46: 739-751. doi: 10.1016/j.plaphy.2008.05.009
|
| [14] |
Zhang BH, Wang QL, Wang KB, et al. (2007) Identification of cotton microRNAs and their targets. Gene 397: 26-37. doi: 10.1016/j.gene.2007.03.020
|
| [15] |
Zhang B, Pan X, Cannon CH, et al. (2006) Conservation and divergence of plant microRNA genes. Plant J 46: 243-259. doi: 10.1111/j.1365-313X.2006.02697.x
|
| [16] |
Frazier TP, Xie F, Freistaedter A, et al. (2010) Identification and characterization of microRNAs and their target genes in tobacco (Nicotiana tabacum). Planta 232: 1289-1308. doi: 10.1007/s00425-010-1255-1
|
| [17] |
Xie F, Frazier T, Zhang B (2010) Identification and characterization of microRNAs and their targets in the bioenergy plant switchgrass (Panicum virgatum). Planta 232: 417-434. doi: 10.1007/s00425-010-1182-1
|
| [18] |
Barozai, MYK, Din M, Baloch IA (2013) Structural and functional based identification of the bean (Phaseolus) microRNAs and their targets from Expressed Sequence Tags. J Struct Funct Genomics 14: 11-18. doi: 10.1007/s10969-013-9152-z
|
| [19] |
Din M, Barozai MYK (2014) Profiling microRNAs and their targets in an important fleshy fruit: Tomato (Solanum lycopersicum). Gene 535: 198-203. doi: 10.1016/j.gene.2013.11.034
|
| [20] | Din M, Barozai MYK (2014) Profiling and characterization of eggplant (Solanum melongena L.) microRNAs and their targets. Mol Biol Rep 41: 889-894. |
| [21] | Din M, Barozai MYK, Baloch IA (2016) Profiling and annotation of microRNAs and their putative target genes in chilli (Capsicum annuum L.) using ESTs. Gene Rep 5: 62-69. |
| [22] |
Muchero W, Diop NN, Bhatetal PR (2009) A consensus genetic map of cowpea (Vigna unguiculata (L) Walp) and synteny based on EST-derived SNPs. Proc Natl Acad Sci U.S.A. 106:18159-18164. doi: 10.1073/pnas.0905886106
|
| [23] |
Pule-Meulenberg F, Belane AK, Krasova-Wade T, et al. (2010) Symbiotic functioning and bradyrhizobial biodiversity of cowpea (Vigna unguiculata L. Walp) in Africa. BMC Microbiol 10: 89. doi: 10.1186/1471-2180-10-89
|
| [24] | Griffiths-Jones S (2004) The microRNA registry. Nucleic Acids Res 32D: 109-111. |
| [25] |
Altschul SF, Gish W, Miller W, et al. (1990) Basic local alignment search tool. J Mol Biol 215: 403-410. doi: 10.1016/S0022-2836(05)80360-2
|
| [26] |
Altschul SF, Madden TL, Schäffer AA, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389-3402. doi: 10.1093/nar/25.17.3389
|
| [27] |
Zuker M (2003) Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res 31: 3406-3415. doi: 10.1093/nar/gkg595
|
| [28] |
Barozai MYK (2012) Identification and characterization of the microRNAs and their targets in Salmo salar. Gene 499: 163-168. doi: 10.1016/j.gene.2012.03.006
|
| [29] | Ambros V, Bartel B, Bartel DP, et al. (2003) A uniform system for microRNA annotation. RNA 9: 277-279. |
| [30] |
Dai X, Zhao PX (2011) psRNATarget: a plant small RNA target analysis server. Nucleic Acids Res 39: 155-159. doi: 10.1093/nar/gkq766
|
| [31] |
Barozai MYK (2012) The microRNAs and their targets in the channel catfish (Ictalurus punctatus). Mol Biol Rep 39: 8867-8872. doi: 10.1007/s11033-012-1753-2
|
| [32] |
Kruger J, Rehmsmeier M (2006) RNAhybrid: microRNA target prediction easy, fast and flexible. Nucl Acids Res 34: 451-454. doi: 10.1093/nar/gkj455
|
| [33] | Barozai MYK, Husnain T (2011) Identification of biotic and abiotic stress up-regulated ESTs in Gossypium arboreum. Mol Biol Rep 39: 1011-1018. |
| [34] | Barozai MYK, Wahid AH (2012) In silico identification and characterization of cumulative abiotic stress responding genes in Potato (Solanum tuberosum L.). Pak J Bot 44: 57-69. |
| [35] |
Barozai MYK, Kakar AG, Din M (2012) The relationship between codon usage bias and salt resistant genes in Arabidopsis thaliana and Oryza sativa. Pure Appl Biol 1: 48-51. doi: 10.19045/bspab.2012.12005
|
| [36] | Barozai MYK, Kakar S, Sarangzai AM (2013) Profiling the carrot (Daucus carota L.) microRNAs and their targets. Pak J Bot 45: 353-358. |
| [37] |
Wang J, Yang X, Xu H, et al. (2012) Identification and characterization of microRNAs and their target genes in Brassica oleracea. Gene 505: 300-308. doi: 10.1016/j.gene.2012.06.002
|
| [38] | Barozai MYK (2013) Identification of microRNAs and their targets in Artemisia annua L. Pak J Bot 45: 461-465. |
| [39] |
Ghani A, Din M, Baloch IA, et al. (2013) Identification of MicroRNA in 12 plant species of fabaceae. Pure Appl Bio 2: 104-115. doi: 10.19045/bspab.2013.23005
|
| [40] | Orlov YL, Dobrovolskaya O, Yuan CH, et al. (2012). Integrative computer analysis of antisense transcripts and miRNA targets in plant genomes. J Stress Physiol Biochem 8: S7. |
| [41] |
Barozai MYK (2012) The novel 172 sheep (Ovis aries) microRNAs and their targets. Mol Biol Rep 39: 6259-6266. doi: 10.1007/s11033-012-1446-x
|
| [42] |
Chen L, Ren YY, Zhang YY, et al. (2012) Genome-wide identification and expression analysis of heat-responsive and novel microRNAs in Populus tomentosa. Gene 504: 160-165. doi: 10.1016/j.gene.2012.05.034
|
| [43] |
Ji Z, Wang G, Xie Z, et al. (2012) Identification and characterization of microRNA in the dairy goat (Capra hircus) mammary gland by Solexa deep sequencing technology. Mol Biol Rep 39: 9361-9371. doi: 10.1007/s11033-012-1779-5
|
| [44] |
Barozai MYK (2012) The microRNAs and their targets in the channel catfish (Ictalurus punctatus). Mol Biol Rep 39: 8867-8872. doi: 10.1007/s11033-012-1753-2
|
| [45] |
Yu J, Wang F, Yang GH, et al. (2006). Human microRNA clusters: genomic organization and expression profile in leukemia cell lines. Biochem Biophys Res Commun 349: 59-68. doi: 10.1016/j.bbrc.2006.07.207
|
| [46] |
Jones-Rhoades MW, Bartel DP (2004) Computational identification of plant microRNAs and their targets, including a stress induced miRNA. Mol Cell 14: 787-799. doi: 10.1016/j.molcel.2004.05.027
|
| [47] |
Crooks GE, Hon G, Chandonia JM, et al. (2004) Web-Logo: a sequence logo generator. Genome Res 14:1188-1190. doi: 10.1101/gr.849004
|
| [48] |
Larkin MA, Blackshields G, Brown NP, et al. (2007) ClustalW and ClustalX version 2. Bioinform 23: 2947-2948. doi: 10.1093/bioinformatics/btm404
|
| [49] | Zeng CY, Wang WQ, Zheng Y, et al.(2009) Conservation and divergence of microRNAs and their functions in Euphorbiaceous plants. Nucleic Acids Res 38: 981-995. |
| [50] |
Bartel DP (2009) MicroRNAs: target recognition and regulatory functions. Cell 136: 215-233. doi: 10.1016/j.cell.2009.01.002
|
| [51] | Kohli P, Kalia M, Gupta R (2015) Pectin Methylesterases: A Review. J Bioprocess Biotech 5: 228. |
| [52] |
Whitney SM, Andrews TJ (2001) The gene for the ribulose-1, 5-bisphosphate carboxylase/oxygenase (Rubisco) small subunit relocated to the plastid genome of tobacco directs the synthesis of small subunits that assemble into Rubisco. Plant Cell 13: 193-205. doi: 10.1105/tpc.13.1.193
|
| [53] |
Ballester AR, Molthoff J, de Vos R, et al. (2010) Biochemical and molecular analysis of pink tomatoes: deregulated expression of the gene encoding transcription factor SlMYB12 leads to pink tomato fruit color. Plant Physiol 152: 71-84. doi: 10.1104/pp.109.147322
|
| [54] |
Kodaira KS, Qin F, Tran LS, et al. (2011) Arabidopsis Cys2/His2 zinc-finger proteins AZF1 and AZF2 negatively regulate abscisic acid-repressive and auxin-inducible genes under abiotic stress conditions. Plant Physiol 157: 742-756. doi: 10.1104/pp.111.182683
|
| [55] |
Soria-Guerra RE, Rosales-Mendoza S, Gasic K, et al. (2011) Gene expression is highly regulated in early developing fruit of apple. Plant Mol Biol Rep 29: 885-897. doi: 10.1007/s11105-011-0300-y
|
| [56] |
Yadav SK (2010) Cold stress tolerance mechanisms in plants. A review. Agron Sustain Dev 30: 515-527. doi: 10.1051/agro/2009050
|
| [57] | Prasad PVV, Staggenborg SA, (2008) Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants, In Ristic, Z. Author, Response of Crops to Limited Water. Madison, WI, USA, 301-355. |
| [58] | Qados AMSA (2011) Effect of salt stress on plant growth and metabolism of bean plant Viciafaba (L.). J Saudi Soc Agric Sci 10: 7-15. |
| [59] |
Rejeb IB, Pastor V, Mauch-Mani B (2014) Plant Responses to Simultaneous Biotic and Abiotic Stress: Molecular Mechanisms. Plants 3:458-475. doi: 10.3390/plants3040458
|
| [60] | Sheshadri SA, Nishanth MJ, Simon B (2016) Stress-mediated cis-element transcription factor interactions interconnecting primary and specialized metabolism in planta. Front Plant Sci 7: 1725. |
| [61] | Fluhr R (2001) Sentinels of disease. Plant resistance genes. Plant Physiol 127: 1367-1374. |
| [62] |
Umezawa T, Yoshida R, Maruyama K, et al. (2004) SRK2C, a SNF1-related protein kinase 2, improves drought tolerance by controlling stress-responsive gene expression in Arabidopsis thaliana. Proc Natl Acad Sci U.S.A. 101: 17306-17311. doi: 10.1073/pnas.0407758101
|
| [63] |
Narayan A, Sachdeva P, Sharma K, et al. (2007) Serine threonine protein kinases of mycobacterial genus: phylogeny to function. Physiol genomics 29: 66-75. doi: 10.1152/physiolgenomics.00221.2006
|
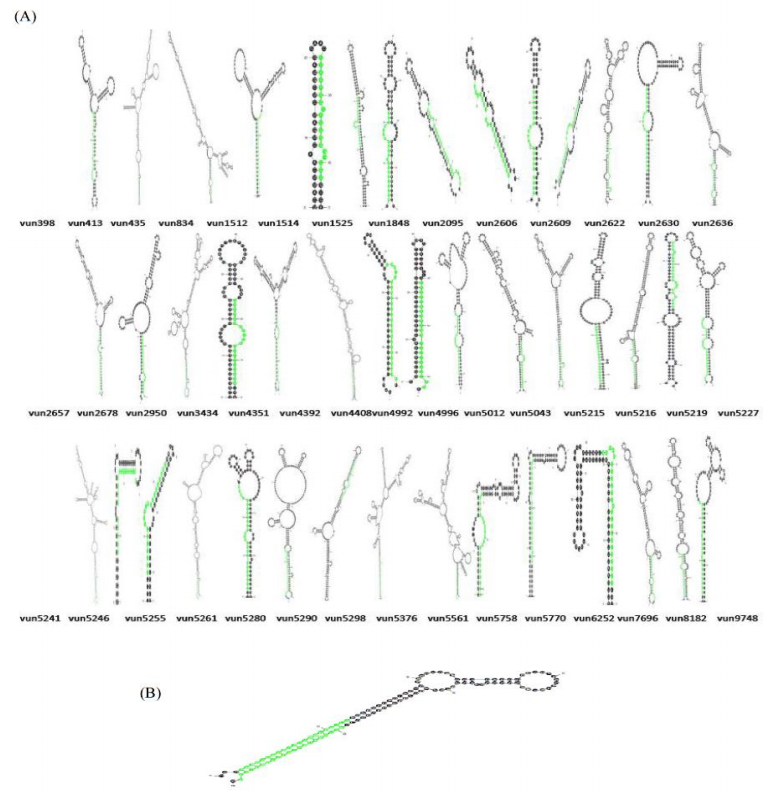
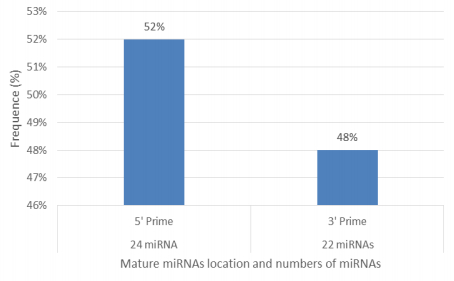
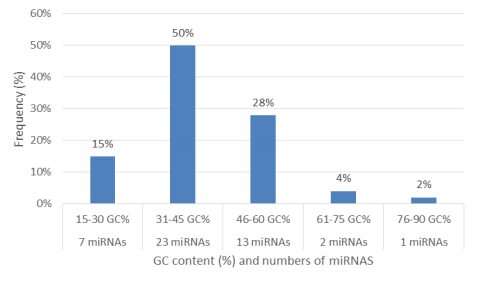
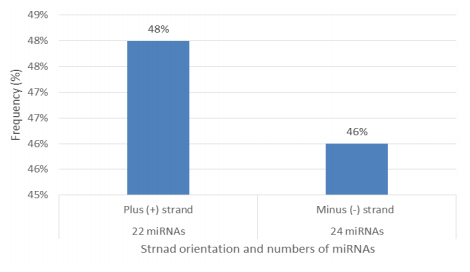
Figures(11) / Tables(2)

Zareen Gul, Muhammad Younas Khan Barozai, Muhammad Din. In-silico based identification and functional analyses of miRNAs and their targets in Cowpea (Vigna unguiculata L.)[J]. AIMS Genetics, 2017, 4(2): 138-165. doi: 10.3934/genet.2017.2.138







 DownLoad:
DownLoad: