Citation: Lewis Trotter, Ashraf Dewan, Todd Robinson. Effects of rapid urbanisation on the urban thermal environment between 1990 and 2011 in Dhaka Megacity, Bangladesh[J]. AIMS Environmental Science, 2017, 4(1): 145-167. doi: 10.3934/environsci.2017.1.145

| [1] |
Grimm NB, Faeth SH, Golubiewski NE, et al. (2008) Global change and the ecology of cities. Science 319: 756-760. doi: 10.1126/science.1150195

|
| [2] | United Nations (UN), World urbanization prospects: the 2011 revision. United Nations, 2012. Available from: http://www.un.org/en/desa/population/publications/pdf/urbanization/WUP2011_Report.pdf |
| [3] | United Nations Population Fund (UNFPA), The state of world population: 2011. United Nations Population Fund, 2011. Available from: http://www.unfpa.org/sites/default/files/pub-pdf/EN-SWOP2011-FINAL.pdf. |
| [4] |
Black D, Henderson V (1999) A theory of urban growth. JPE 107: 252-284. doi: 10.1086/250060
|
| [5] |
Dewan AM, Yamaguchi Y (2009) Land use and land cover change in Greater Dhaka, Bangladesh: Using remote sensing to promote sustainable urbanization. Appl Geogr 29: 390-401. doi: 10.1016/j.apgeog.2008.12.005
|
| [6] |
Rana MMP (2011) Urbanization and sustainability: challenges and strategies for sustainable urban development in Bangladesh. Environment, Development and Sustainability 13: 237-256. doi: 10.1007/s10668-010-9258-4
|
| [7] |
Hasan S, Mulamoottil G (1994) Environmental problems of Dhaka City: a study of mismanagement. Cities 11: 195-200. doi: 10.1016/0264-2751(94)90059-0
|
| [8] |
Azad A, Kitada T (1998) Characteristics of the air pollution in the city of Dhaka, Bangladesh in winter. Atmospheric Environment 32: 1991-2005. doi: 10.1016/S1352-2310(97)00508-6
|
| [9] | Dewan AM, Corner RJ (2013) Introduction to Dhaka Megacity, In: Dewan A. M & Corner R. J, Dhaka megacity: geospatial perspectives on urbanisation, environment and health, 2 Eds., New York: Springer Science & Business Media, 1-48. |
| [10] |
Foley JA, Defries R, Asner GP, et al. (2005) Global consequences of land use. Science 309: 570-574. doi: 10.1126/science.1111772
|
| [11] |
Voogt JA, Oke TR (1998) Effects of urban surface geometry on remotely-sensed surface temperature. Int J Remote Sens 19: 895-920. doi: 10.1080/014311698215784
|
| [12] |
Yuan F, Bauer ME (2007) Comparison of impervious surface area and normalized difference vegetation index as indicators of surface urban heat island effects in Landsat imagery. Remote Sens Environ 106: 375-386. doi: 10.1016/j.rse.2006.09.003
|
| [13] |
Kovats S, Akhtar H (2008) Climate, climate change and human health in Asian cities. Environ Urban 20: 165-175. doi: 10.1177/0956247808089154
|
| [14] |
Patz JA, Olson SH (2006) Climate change and health: global to local influences on disease risk. Ann Trop Med Parasit 100: 535-549. doi: 10.1179/136485906X97426
|
| [15] | Shahid S (2009) Probable impacts of climate change on public health in Bangladesh. Asia Pac J Public Health 124: 432-444. |
| [16] |
Hashizume M, Dewan AM, Sunahara T, et al. (2012) Hydroclimatological variability and dengue transmission in Dhaka, Bangladesh: a time-series study. BMC Infect Dis 12: 98. doi: 10.1186/1471-2334-12-98
|
| [17] | Dewan AM, Corner R, Hashizume M, et al. (2014) Typhoid fever and its association with environmental factors in the Dhaka metropolitan area of Bangladesh: a spatial and time-series approach. PLoS Negl Trop Dis 7: 1998. |
| [18] | Byomkesh T, Nakagoshi N, Dewan AM (2011) Urbanization and green space dynamics in greater Dhaka, Bangladesh. Landsc Ecol Eng 8: 45-58. |
| [19] | Fortuniak K, Kłysik K, Wibig J (2005) Urban–rural contrasts of meteorological parameters in Łódź. Theor Appl Climatol 84: 91-101. |
| [20] |
Wong NH, Yu C (2005) Study of green areas and urban heat island in a tropical city. Habitat Int 29: 547-558. doi: 10.1016/j.habitatint.2004.04.008
|
| [21] |
Saaroni H, Ben-Dor E, Bitan A, et al. (2000) Spatial distribution and microscale characteristics of the urban heat island in Tel-Aviv, Israel. Landscape Urban Plan 48: 1-18. doi: 10.1016/S0169-2046(99)00075-4
|
| [22] |
Yow DM (2007) Urban heat islands: observations, impacts and adaptation. Geography Compass 1: 1227-1251. doi: 10.1111/j.1749-8198.2007.00063.x
|
| [23] |
Weng Q (2009) Thermal infrared remote sensing for urban climate and environmental studies: methods, applications and trends. ISPRS J Photogramm 64: 335-344. doi: 10.1016/j.isprsjprs.2009.03.007
|
| [24] |
Zhou X, Wang YC (2011) Dynamics of land surface temperature in response to land-use/cover change. Geogr Res 49: 23-36. doi: 10.1111/j.1745-5871.2010.00686.x
|
| [25] |
Carnahan WH, Larson RC (1990) An analysis of an urban heat sink. Remote Sens Environ 33: 65-71. doi: 10.1016/0034-4257(90)90056-R
|
| [26] |
Streutker DR (2002) A remote sensing study of the urban heat island of Houston, Texas. Int J Remote Sens 23: 2595-2608. doi: 10.1080/01431160110115023
|
| [27] | Walawender JP, Szymanowski M, Hajto MJ, et al. (2013) Land surface temperature patterns in the urban agglomeration of Krakow (Poland) derived from landsat-7/etm+ data. Pure Appl Geophys 4: 23-54. |
| [28] |
Voogt JA, Oke TR (2003) Thermal remote sensing of urban climates. Remote Sens Environ 86: 370-384. doi: 10.1016/S0034-4257(03)00079-8
|
| [29] |
Carlson TN, Gillies RR, Perry EM (1994) A method to make use of thermal infrared temperature and NDVI measurements to infer surface soil water content and fractional vegetation cover. Remote Sens Rev 9: 161-173. doi: 10.1080/02757259409532220
|
| [30] |
Zha Y, Gao J, Ni S (2003) Use of normalized difference built-up index in automatically mapping urban areas from TM imagery. Int J Remote Sens 24: 583-594. doi: 10.1080/01431160304987
|
| [31] |
Weng Q (2008) Medium spatial resolution satellite imagery for estimating and mapping urban impervious surfaces using LSMA and ANN. IEEE T Geosci Remote 46: 2397-2406. doi: 10.1109/TGRS.2008.917601
|
| [32] |
Ma Y, Kuang Y, Huang N (2010) Coupling urbanization analyses for studying urban thermal environment and its interplay with biophysical parameters based on TM/ETM+ imagery. Int J Appl Earth Obs 12: 110-118. doi: 10.1016/j.jag.2009.12.002
|
| [33] |
Adams J (1995) Classification of multispectral images based on fractions of endmembers: application to land-cover change in the Brazilian Amazon. Remote Sens Environ 52: 137-154. doi: 10.1016/0034-4257(94)00098-8
|
| [34] |
Brown M, Lewis HG, Gunn SR (2000) Linear spectral mixture models and support vector machines for remote sensing. IEEE T Geosci Remote 38: 2346-2360. doi: 10.1109/36.868891
|
| [35] | Rashid H (1978) Geography of Dhaka, In: Rashid, H, Geography of Bangladesh, 2 Eds., Dhaka: University Press, 78-94. |
| [36] | Tareq SM, Maruo M, Ohta K (2013) Characteristics and role of groundwater dissolved organic matter on arsenic mobilization and poisoning in Bangladesh. Phys Chem Earth 1: 77-84. |
| [37] | Water Resources Planning Organization (WARPO), Datum and map projections for GIS and GPS applications in Bangladesh. Water Resources Planning Organization, 1996. Available from: http://www.cegisbd.com/pdf/tn10DatumMapProjections.pdf. |
| [38] |
Chander G, Markham B (2003) Revised landsat-5 TM radiometric calibration procedures and postcalibration dynamic ranges. IEEE T Geosci Remote 41: 2674-2677. doi: 10.1109/TGRS.2003.818464
|
| [39] |
Chander G, Markham BL, Helder DL (2009) Summary of current radiometric calibration coefficients for Landsat MSS, TM, ETM+ and EO-1 ALI sensors. Remote Sens Environ 113: 893-903. doi: 10.1016/j.rse.2009.01.007
|
| [40] | Chavez P (1996) Image-based atmospheric corrections: revisited and improved. Photogramm eng rem s 62: 1025-1035. |
| [41] | IDRISI Selva-GIS and Image Processing Software (Version 17), (2012). Worcester, Massachusetts: Clark Laboratories. |
| [42] | Anderson R, Hardy EE, Roach JT, et al. (1976) A land use and land cover classification system for use with remote sensor data, In: United States Geological Survey (USGS), Professional Papers Vol. 964, 1 Eds., Sioux Falls: USGS Professional Printing, 41-78. |
| [43] |
Mas J (1999) Monitoring land-cover changes: a comparison of change detection techniques. Int J Remote Sens 20: 139-152. doi: 10.1080/014311699213659
|
| [44] | Streatfield P, Karar Z (2008) Population challenges for Bangladesh in the coming decades. JHPS 26: 261-272. |
| [45] |
Qin Z, Karnieli A, Berliner P (2001) A mono-window algorithm for retrieving land surface temperature from Landsat TM data and its application to the Israel-Egypt border region. Int J Remote Sens 22: 3719-3746. doi: 10.1080/01431160010006971
|
| [46] |
Jiménez-Muñoz JC, Sobrino JA (2003) A generalised single channel method for retrieving land surface temperature from remote sensing data. J Geophys Res 108: 4688. doi: 10.1029/2003JD003480
|
| [47] |
Chen XL, Zhao HM, Li P, et al. (2006) Remote sensing image-based analysis of the relationship between urban heat island and land use/cover changes. Remote Sens Environ 104: 133-146. doi: 10.1016/j.rse.2005.11.016
|
| [48] |
Lo CP, Quattrochi DA (2003) Land-use and land-cover change, urban heat island phenomenon, and health implications. Photogramm Eng Rem S 69: 1053-1063. doi: 10.14358/PERS.69.9.1053
|
| [49] | Tran H, Uchihama D, Ochi S, et al. (2006) Assessment with satellite data of the urban heat island effects in Asian mega cities. Int J Appl Earth Obs 8: 147-156. |
| [50] |
Artis DA, Carnahan WH (1982) Survey of emissivity variability in thermography of urban areas. Remote Sens Environ 12: 313-329. doi: 10.1016/0034-4257(82)90043-8
|
| [51] |
Sobrino JA, Jiménez-Muñoz JC, Sòria G, et al. (2008) Land surface emissivity retrieval from different VNIR and TIR sensors. Int J Remote Sens 46: 316-327. doi: 10.1109/TGRS.2007.904834
|
| [52] |
Xiao R, Ouyang Z, Zheng H, et al. (2007) Spatial pattern of impervious surfaces and their impacts on land surface temperature in Beijing, China. J Environ Sci 19: 250-256. doi: 10.1016/S1001-0742(07)60041-2
|
| [53] |
Mackey CW, Lee X, Smith RB (2012) Remotely sensing the cooling effects of city scale efforts to reduce urban heat island. Build Environ 49: 348-358. doi: 10.1016/j.buildenv.2011.08.004
|
| [54] | Nichol JE (1994) Approach to a GIS-Based monitoring survey of microclimate of Singapore's housing estates. Cities 60: 43-56. |
| [55] |
Sobrino JA, Jiménez-Muñoz JC, Paolini L (2004) Land surface temperature retrieval from Landsat TM 5. Remote Sens Environ 90: 434-440. doi: 10.1016/j.rse.2004.02.003
|
| [56] |
Carlson TN, Ripley DA (1997) On the relation between NDVI, fractional vegetation cover and leaf area index. Remote Sens Environ 62: 241-252. doi: 10.1016/S0034-4257(97)00104-1
|
| [57] |
Sobrino JA, Caselles V, Becker F (1990) Significance of the remotely sensed thermal infrared measurements obtained over a citrus orchard. ISPRS J Photogramm 44: 343-354. doi: 10.1016/0924-2716(90)90077-O
|
| [58] | Walawender JP, Hajto MJ, Iwaniuk P (2012) A new ArcGIS toolset for automated mapping of land surface temperature with the use of landsat satellite data. IEEE International Geoscience and Remote Sensing Symposium 2012: 4371-4374. |
| [59] |
Carlson TN, Arthur ST (2000) The impact of land use–land cover changes due to urbanization on surface microclimate and hydrology: a satellite perspective. Global Planet Change 25: 49-65. doi: 10.1016/S0921-8181(00)00021-7
|
| [60] |
Weng Q, Lu D, Schubring J (2004) Estimation of land surface temperature–vegetation abundance relationship for urban heat island studies. Remote Sens Environ 89: 467-483. doi: 10.1016/j.rse.2003.11.005
|
| [61] |
Zhang X, Zhong T, Wang K, et al. (2009) Scaling of impervious surface area and vegetation as indicators to urban land surface temperature using satellite data. Int J Remote Sens 30: 841-859. doi: 10.1080/01431160802395219
|
| [62] |
Huete A, Didan K, Miura T, et al. (2002) Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens Environ 83: 195-213. doi: 10.1016/S0034-4257(02)00096-2
|
| [63] |
Sun F, Sun W, Chen J, et al. (2012). Comparison and improvement of methods for identifying waterbodies in remotely sensed imagery. Int J Remote Sens 33: 6854-6875. doi: 10.1080/01431161.2012.692829
|
| [64] | ArcGIS-GIS Software (Version 10.1), (2012). Redlands, California: Environmental Systems Research Institute (ESRI). |
| [65] |
Yang X, Liu Z (2005) Use of satellite-derived landscape imperviousness index to characterize urban spatial growth. Computers, Environment and Urban Systems 29: 524-540. doi: 10.1016/j.compenvurbsys.2005.01.005
|
| [66] | ENVI-GIS software (Version 4.8), (2012). Boulder, Colorado: Harris Geospatial Solutions. |
| [67] |
Wu C (2004) Normalized spectral mixture analysis for monitoring urban composition using ETM+ imagery. Remote Sens Environ 93: 480-492. doi: 10.1016/j.rse.2004.08.003
|
| [68] |
Green AA, Berman M, Switzer P, et al. (1988) A transformation for ordering multispectral data in terms of image quality with implications for noise removal. IEEE T Geosci Remote 26: 65-74. doi: 10.1109/36.3001
|
| [69] | Boardman J (1993) Automating spectral unmixing of AVIRIS data using convex geometry concepts. 4th Annual JPL Airborne Geoscience Conference and Worskhop 2: 2-5. |
| [70] | Boardman J, Kruse F, Green R (1995) Mapping target signatures via partial unmixing of AVIRIS data. 5th Annual JPL Airborne Geoscience Conference and Worskhop 3: 3-6. |
| [71] | Tukey JW (1953) The problem of multiple comparisons. In: Unpublished Manuscript, Trenton: Princeton University, 1-38. |
| [72] |
Liu H, Weng Q (2008) Seasonal variations in the relationship between landscape pattern and land surface temperature in Indianapolis, USA. Environ Monit Assess 144: 199-219. doi: 10.1007/s10661-007-9979-5
|
| [73] | National Aeronautics and Space Administration, Landsat 7 science data users handbook. National Aeronautics and Space Administration, 2000. Available from: https://landsat.gsfc.nasa.gov/wp-content/uploads/2016/08/Landsat7_Handbook.pdf. |
| [74] | SPSS- Statistics Software (Version 22), (2012). Armonk, New York: IBM Corporation. |
| [75] | Cohen J, Cohen P, West S, et al. (2013) Data-Analytic Strategies Using Multiple Regression/Correlation, In: Cohen J, Cohen P, Applied multiple regression/correlation analysis for the behavioural sciences, 3 Eds., London: Lawrence Erlbaum Associates, 151-192. |
| [76] |
Ahmed B, Kamruzzaman M, Zhu X, et al. (2013) Simulating land cover changes and their impacts on land surface temperature in Dhaka, Bangladesh. Remote Sens 5: 5969-5998. doi: 10.3390/rs5115969
|
| [77] | Raja DR (2012) Spatial analysis of land surface temperature in Dhaka metropolitan area. J Bangladesh Institute of Planners 5: 151-167. |
| [78] |
Xiao H, Weng Q (2007) The impact of land use and land cover changes on land surface temperature in a karst area of China. J Environ Manage 85: 245-257. doi: 10.1016/j.jenvman.2006.07.016
|
| [79] |
Xu H, Dongfeng L, Tang F (2013) The impact of impervious surface development on land surface temperature in a subtropical city: Xiamen, China. Int J Climatol 33: 1873-1883. doi: 10.1002/joc.3554
|
| [80] |
Li Y, Zhang H, Kainz W (2012) Monitoring patterns of urban heat islands of the fast-growing Shanghai metropolis, China: Using time-series of Landsat TM/ETM+ data. Int J Appl Earth Obs 19: 127-138. doi: 10.1016/j.jag.2012.05.001
|
| [81] |
Tan KC, San-Lim H, Matjafri MZ, et al. (2010) Landsat data to evaluate urban expansion and determine land use/land cover changes in Penang Island, Malaysia. Environ Earth Sci 60: 1509-1521. doi: 10.1007/s12665-009-0286-z
|
| [82] |
Li J, Wang J, Ma J, et al. (2009) Remote sensing evaluation of urban heat island and its spatial pattern of the Shanghai metropolitan area, China. Ecol Complex 6: 413-420. doi: 10.1016/j.ecocom.2009.02.002
|
| [83] |
Li J, Song C, Cao L, et al. (2011) Impacts of landscape structure on surface urban heat islands: a case study of Shanghai, China. Remote Sens Environ 115: 3249-3263. doi: 10.1016/j.rse.2011.07.008
|
| [84] |
Roth M, Oke TR, Emery WJ (1989) Satellite-derived urban heat islands from three coastal cities and the utilization of such data in urban climatology. Int J Remote Sens 10: 1699-1720. doi: 10.1080/01431168908904002
|
| [85] |
Owen TW, Carlson TN, Gillies RR (1998) An assessment of satellite remotely-sensed land cover parameters in quantitatively describing the climatic effect of urbanization. Int J Remote Sens 19: 1663-1681. doi: 10.1080/014311698215171
|
| [86] | Cao L, Li P, Zhang L, et al. (2002) Remote sensing image-based analysis of the relationship between urban heat island and vegetation fraction. The International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences 1: 1379-1383. |
| [87] |
Giri C, Pengra B, Zhu Z, et al. (2007) Monitoring mangrove forest dynamics of the sundarbans in Bangladesh and India using multi-temporal satellite data from 1973 to 2000. Estuar Coast Shelf S 73: 91-100. doi: 10.1016/j.ecss.2006.12.019
|
| [88] | Bangladesh Department of Environment Climate Change Cell (2006) Bangladesh climate change impacts and vulnerability: a synthesis. Dhaka, Department of Environment Publishing. |
| [89] | Goward SN (1981) Thermal behavior of urban landscapes and the urban heat island. Phys Geogr 2: 19-33. |
| [90] | Grimmond CSB (2005) Progress in measuring and observing the urban atmosphere. Theor Appl Climatol 84: 3-22. |
| [91] | Oke TR (1982) The energetic basis of the urban heat island. J Roy Meteor Soc 108: 1-24. |
| [92] | Xu H, Ding F, Wen X (2009) Urban expansion and heat island dynamics in the Quanzhou region, China. IEEE J-STAEORS 2: 74-79. |
| [93] | Weng Q (2001) A remote sensing GIS evaluation of urban expansion and its impact on surface temperature in the Zhujiang Delta, China. Int J Remote Sens 22: 1999-2014. |
| [94] | Dewan A, Corner R (2014). Impact of land use and land cover changes on urban land surface temperature, In: Dewan AM, Corner RJ, Dhaka megacity: geospatial perspectives on urbanisation, environment and health, 2 Eds., New York: Springer Science & Business Media, 219-238. |
| [95] | Raja DR, Neema MN (2013) Impact of urban development and vegetation on land surface temperature of Dhaka city. Comp Sci Appl 7973: 351-367. |
| [96] |
Weng Q, Hu X, Lu D (2008). Extracting impervious surfaces from medium spatial resolution multispectral and hyperspectral imagery: a comparison. Int J Remote Sens 29: 3209-3232. doi: 10.1080/01431160701469024
|
| [97] |
Song C, Woodcock CE, Seto KC (2001) Classification and change detection using landsat TM data. Remote Sens Environ 75: 230-244. doi: 10.1016/S0034-4257(00)00169-3
|
| [98] |
Weng Q, Lu D (2008) Extracting impervious surfaces from medium spatial resolution multispectral and hyperspectral imagery: a comparison. Int J Remote Sens 29: 3209-3232. doi: 10.1080/01431160701469024
|
| [99] | Xu H (2008) A new remote sensing index for fastly extracting impervious surface information. GIS 8: 12-28. |

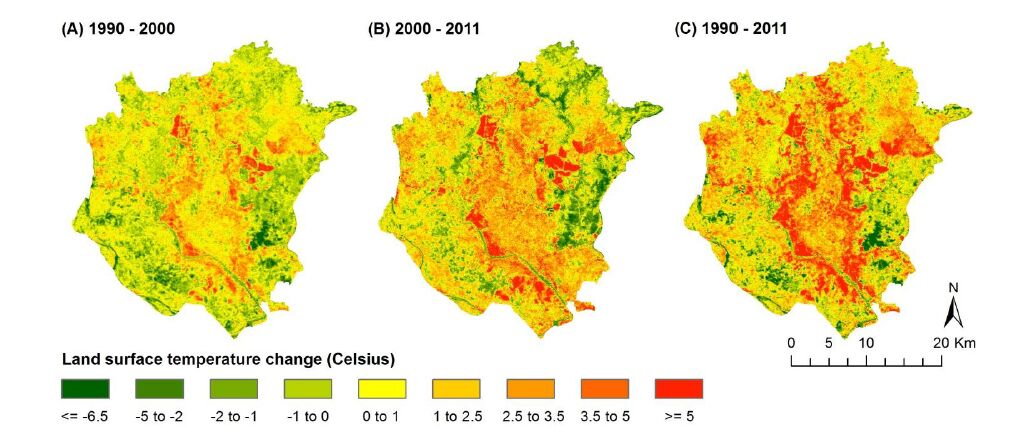
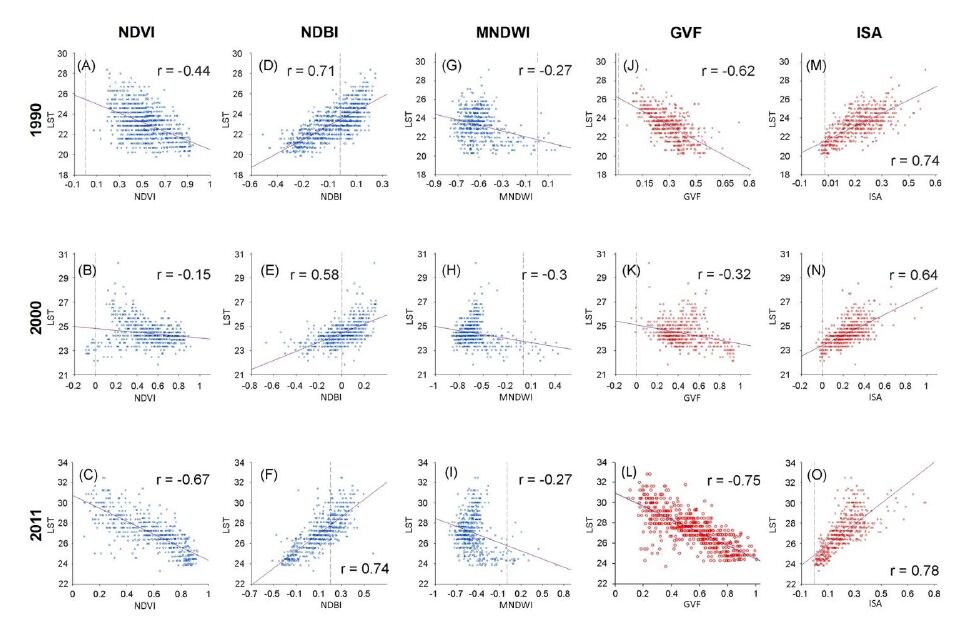
Figures(7) / Tables(4)

Lewis Trotter, Ashraf Dewan, Todd Robinson. Effects of rapid urbanisation on the urban thermal environment between 1990 and 2011 in Dhaka Megacity, Bangladesh[J]. AIMS Environmental Science, 2017, 4(1): 145-167. doi: 10.3934/environsci.2017.1.145







 DownLoad:
DownLoad: