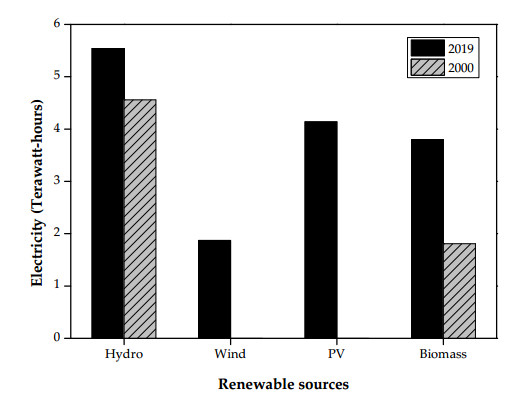
In response to the international agreement on the climate change and the energy supply diversification, the Taiwan government actively promoted the use of wind power since 2000. This paper highlighted the Taiwan's wind resources, and interactively analyzed the regulatory strategies for promoting wind power and trend analysis of wind power over the two past decades (2000–2019). In the first development stage, the electricity generation by onshore wind power rapidly grew from 1.38 gigawatt-hours (GW-h) in 2000 to 1,492.7 GW-h in 2011 because of the implementations of promotional measures and official subsidies. However, its growth rate became slow due to the restrictions on available land for the onshore wind power farms. With the promulgation of the Renewable Energy Development Act in 2009, the government not only declared preferable feed-in-tariff (FIT) rates, but also announced some incentive and promotional plans, focusing on offshore wind power farms from the early 2010s. Therefore, the electricity generation by offshore wind power significantly increased from 26.7 GW-h in 2018 to 175.4 GW-h in 2019. Using the data on the electricity generation by wind power in 2019 (i.e., 1,871.3 × 106 kW-h) and the average default value (i.e., 0.539 kg CO2/kW-h), the equivalent mitigation of CO2 emission from the wind power systems in Taiwan was thus estimated to be about one million metric tons. Based on the progressive development of wind power and other renewable resources over the past two decades, one of the sustainable development goals (SDGs) for nuclear-free homeland and 20% renewable electricity generation may be prospective in the energy transition by 2025. However, this goal has faced serious challenges because the corona virus disease 2019 (COVID-19) has caused the delays in the Taiwanese wind power industry.
Citation: Wen-Tien Tsai. Overview of wind power development over the two past decades (2000-2019) and its role in the Taiwan's energy transition and sustainable development goals[J]. AIMS Energy, 2021, 9(2): 342-354. doi: 10.3934/energy.2021018
In response to the international agreement on the climate change and the energy supply diversification, the Taiwan government actively promoted the use of wind power since 2000. This paper highlighted the Taiwan's wind resources, and interactively analyzed the regulatory strategies for promoting wind power and trend analysis of wind power over the two past decades (2000–2019). In the first development stage, the electricity generation by onshore wind power rapidly grew from 1.38 gigawatt-hours (GW-h) in 2000 to 1,492.7 GW-h in 2011 because of the implementations of promotional measures and official subsidies. However, its growth rate became slow due to the restrictions on available land for the onshore wind power farms. With the promulgation of the Renewable Energy Development Act in 2009, the government not only declared preferable feed-in-tariff (FIT) rates, but also announced some incentive and promotional plans, focusing on offshore wind power farms from the early 2010s. Therefore, the electricity generation by offshore wind power significantly increased from 26.7 GW-h in 2018 to 175.4 GW-h in 2019. Using the data on the electricity generation by wind power in 2019 (i.e., 1,871.3 × 106 kW-h) and the average default value (i.e., 0.539 kg CO2/kW-h), the equivalent mitigation of CO2 emission from the wind power systems in Taiwan was thus estimated to be about one million metric tons. Based on the progressive development of wind power and other renewable resources over the past two decades, one of the sustainable development goals (SDGs) for nuclear-free homeland and 20% renewable electricity generation may be prospective in the energy transition by 2025. However, this goal has faced serious challenges because the corona virus disease 2019 (COVID-19) has caused the delays in the Taiwanese wind power industry.

| [1] | World Economic Outlook Databases (2019). Available from: https://www.imf.org/external/pubs/ft/weo/2019/02/weodata/index.aspx. |
| [2] | Ministry of Economic Affairs (2020). Energy statistics handbook-2019. MOEA: Taipei (Taiwan). |
| [3] |
Wu JH, Huang YH (2006) Renewable energy perspectives and support mechanisms in Taiwan. Renew Energy 31: 1718-1732. doi: 10.1016/j.renene.2005.09.007

|
| [4] |
Chen FL, Lu SM, Chang YL (2007) Renewable energy in Taiwan: Its developing status and strategy. Energy 32: 1634-1646. doi: 10.1016/j.energy.2006.12.007
|
| [5] |
Hwang JJ (2010) Promotional policy for renewable energy development in Taiwan. Renew Sustain Energy Rev 14: 1079-1087. doi: 10.1016/j.rser.2009.10.029
|
| [6] |
Liou HM (2010) Policies and legislation driving Taiwan's development of renewable energy. Renew Sustain Energy Rev 14: 1763-1781. doi: 10.1016/j.rser.2010.02.013
|
| [7] |
Chen HH, Lee AHI (2014) Comprehensive overview of renewable energy development in Taiwan. Renew Sustain Energy Rev 37: 215-228. doi: 10.1016/j.rser.2014.04.055
|
| [8] |
Balat M (2010) Sustainable developments of wind power in Europe. Part 1: utilization and future prospects. Energy Sources B 5: 409-423. doi: 10.1080/15567240802532528
|
| [9] |
Pablo-Romero MP, Pozo-Barajas R (2017) Global changes in total and wind electricity (1990-2014). AIMS Energy 5: 290-312. doi: 10.3934/energy.2017.2.290
|
| [10] |
Soares-Ramos EPP, de Oliveira-Assis L, Sarrias-Mena R, et al. (2020) Current status and future trends of offshore wind power in Europe. Energy 202: 117787. doi: 10.1016/j.energy.2020.117787
|
| [11] |
Dawn S, Tiwari PK, Goswami AK, et al. (2019) Wind power: Existing status, achievements and government's initiative towards renewable power dominating India. Energy Strategy Rev 23: 178-199. doi: 10.1016/j.esr.2019.01.002
|
| [12] |
Zhang S, Wei J, Chen X, et al. (2020) China in global wind power development: Role, status and impact. Renew Sustain Energy Rev 127: 109881. doi: 10.1016/j.rser.2020.109881
|
| [13] | Energy Statistics Information Query System (2020). Available from: https://www.moeaboe.gov.tw/wesnq/. |
| [14] | Guidelines on Energy Development (2017). Available from: https://adapt.epa.gov.tw/TCCIP-1-D/TCCIP-1-D-6. |
| [15] | Laws and Regulation Retrieving System (2021). Available from: https://law.moj.gov.tw/Eng/index.aspx. |
| [16] |
Shieh CL, Wang CM, Chen YS, et al. (2010) An overview of disasters resulted from Typhoon Morakot in Taiwan. J Disaster Res 5: 236-244. doi: 10.20965/jdr.2010.p0236
|
| [17] | Global Offshore Wind Speeds Rankings (2020). Available from: https://www.4coffshore.com/windfarms/windspeeds.aspx. |
| [18] |
Fang HF (2014) Wind energy potential assessment for the offshore areas of Taiwan west coast and Penghu Archipelago. Renew Energy 67: 237-241. doi: 10.1016/j.renene.2013.11.047
|
| [19] |
Chang PC, Yang RY, Lai CM (2015) Potential of offshore wind energy and extreme wind speed forecasting on the west coast of Taiwan. Energies 8: 1685-1700. doi: 10.3390/en8031685
|
| [20] |
Valentin SV (2010) A STEP toward understanding wind power development policy barriers in advanced economies. Renew Sustain Energy Rev 14: 2796-2807. doi: 10.1016/j.rser.2010.07.043
|
| [21] | Lu SM (2017) Renewable Energies in Taiwan: Reserve, Status, Policy; LAP LAMBERT Academic Publishing: Saarbrucken, Germany, 2017. |
| [22] |
Lin CJ, Yu OS, Chang CL, et al. (2007) Challenges of wind farms connection to future power systems in Taiwan. Renew Energy 34: 1926-1930. doi: 10.1016/j.renene.2008.12.004
|
| [23] |
Lee YM, Tzeng YE (2008) Development and life-cycle inventory analysis of wind energy in Taiwan. J Energy Eng 134: 53-57. doi: 10.1061/(ASCE)0733-9402(2008)134:2(53)
|
| [24] |
Yue CD, Yang MH (2009) Exploring the potential of wind energy for a coastal state. Energy Policy 37: 3925-3940. doi: 10.1016/j.enpol.2009.04.055
|
| [25] |
Chen FL, Lu SM, Tseng KT, et al. (2010) Assessment of renewable energy reserves in Taiwan. Renew Sustain Energy Rev 14: 2511-2528. doi: 10.1016/j.rser.2010.06.021
|
| [26] |
Lee TL (2010) Assessment of the potential of offshore wind energy in Taiwan using fuzzy analytic hierarchy process. Open Civil Eng J 4: 96-104. doi: 10.2174/1874149501004010096
|
| [27] |
Chou CJ, Wu YK, Han GY, et al. (2012) Comparative evaluation of the HVDC and HVAC links integrated in a large offshore wind farm-an actual case study in Taiwan. IEEE Trans Ind Appl 48: 1639-1648. doi: 10.1109/TIA.2012.2209622
|
| [28] |
Kao SM, Pearre NS (2017) Administrative arrangement for offshore wind power developments in Taiwan: Challenges and prospects. Energy Policy 109: 463-472. doi: 10.1016/j.enpol.2017.07.027
|
| [29] |
Zhang Y, Zhang C, Chang YC, et al. (2017) Offshore wind farm in marine spatial planning and the stakeholders engagement: Opportunities and challenges for Taiwan. Ocean Coast Manag 149: 69-80. doi: 10.1016/j.ocecoaman.2017.09.014
|
| [30] | Chien KH (2020) Pacing for renewable energy development: The development state in Taiwan's offshore wind power. Ann Am Assoc Geogr 110: 793-807. |
| [31] |
Valentin SV (2010) Disputed wind directions: Reinvigorating wind power development in Taiwan. Energy Sustain Dev 14: 22-34. doi: 10.1016/j.esd.2009.12.005
|
| [32] |
Huang YH, Wu JH (2011) Assessment of the feed-in tariff mechanism for renewable energies in Taiwan. Energy Policy 39: 8106-8115. doi: 10.1016/j.enpol.2011.10.005
|
| [33] |
Tsai WT (2013) Prospects of renewable energy development in Taiwan. Energy Sources B 8: 263-269. doi: 10.1080/15567240903394281
|
| [34] |
Tsai WT (2014) Feed-in tariff promotion and innovative measures for renewable electricity: Taiwan case analysis. Renew Sustain Energy Rev 40: 1126-1132. doi: 10.1016/j.rser.2014.08.019
|
| [35] |
Chang CT, Lee HC (2016) Taiwan's renewable energy strategy and energy-intensive industrial policy. Renew Sustain Energy Rev 64: 456-465. doi: 10.1016/j.rser.2016.06.052
|
| [36] |
Kung CC, Zhang L, Chang MS (2017) Promotion policies for renewable energy and their effects in Taiwan. J Clean Prod 142: 965-975. doi: 10.1016/j.jclepro.2016.05.034
|
| [37] |
Kung CC, McCarl BA (2020) The potential role of renewable electricity generation in Taiwan. Energy Policy 138: 111227. doi: 10.1016/j.enpol.2019.111227
|
| [38] | Electricity carbon emission factor (2020). Available from: https://www.moeaboe.gov.tw/ecw/populace/content/ContentDesc.aspx?menu_id=6989. |
| [39] | World Health Organization (2020). WHO coronavirus disease (COVID-19) dashboard. Available from: https://covid19.who.int/. |
| [40] |
Cheval S, Adamescu CM, Georgiadis T, et al. (2020) Observed and potential impacts of the COVID-19 pandemic on the environment. Int J Environ Res Public Health 17: 4140. doi: 10.3390/ijerph17114140
|
| [41] | International Energy Agency (2020) Covid-19 Impact on Renewable Energy Growth, Available from: https://www.iea.org/reports/renewable-energy-market-update/covid-19-impact-on-renewable-energy-growth. |
| [42] | Eroglu H (2020) Effects of Covid-19 outbreak on environment and renewable energy sector. Environ Dev Sustain (in press). Available from: https://doi.org/10.1007/s10668-020-00837-4. |
| [43] |
Gillingham KT, Knittel CR, Li J, et al. (2020) The Short-run and Long-run Effects of Covid-19 on Energy and the Environment. Joule 4: 1337-1341. doi: 10.1016/j.joule.2020.06.010
|

Figures(2) / Tables(5)

Wen-Tien Tsai. Overview of wind power development over the two past decades (2000-2019) and its role in the Taiwan's energy transition and sustainable development goals[J]. AIMS Energy, 2021, 9(2): 342-354. doi: 10.3934/energy.2021018







 DownLoad:
DownLoad: