Citation: Kushan Tharuka Lulbadda, K. T. M. U. Hemapala. The additional functions of smart inverters[J]. AIMS Energy, 2019, 7(6): 971-988. doi: 10.3934/energy.2019.6.971

| [1] | From Smart Meters to Smart Inverters: How DVI Is Optimizing the Distributed Grid Greentechmedia.com, 2018. Available from: https://www.greentechmedia.com/articles/read/DVIs-grid-edge-vision-from-smart-meters-to-smart-inverters. |
| [2] | Enayati B (2018) Impacts of the IEEE 1547, standard of DER interconnections, on smart inverters. National Grid: 1-13. |
| [3] | Liu YJ, Lan PH, Lin HH (2014) Grid-connected PV inverter test system for solar photovoltaic power system certification. IEEE Power Energy Soc Gen Meet: 1-5. |
| [4] | Peng W. General Application of Smart inverters in Distribution and Smart Grid. University of Nevada, Las Vegas, August 2015. |
| [5] | De Muro AG, Rodriguez-Seco JE, Zabala E, et al. (2009) Inverter interconnection tests performed in the LABEIN-Tecnalia microgrid involved in the DERlab round-robin testing activity. 2009 10th International Conference on Electrical Power Quality and Utilisation, Lodz, Poland. |
| [6] | Jamal H, Albatran S, Smadi IA (2016) Optimal design of output LC filter and cooling for three-phase voltage-source inverters using teaching-learning-based optimization. 2016 IEEE Energy Conversion Congress and Exposition (ECCE), Milwaukee, USA. |
| [7] |
Formica TJ, Khan HA, Pecht MG (2017) The effect of inverter failures on the return on investment of solar photovoltaic systems. IEEE Access 5: 21336-21343. doi: 10.1109/ACCESS.2017.2753246

|
| [8] | Kabalci E (2017) A smart monitoring infrastructure design for distributed renewable energy systems. Energ convers manage 90: 336-346. |
| [9] | Moreno-García IM, Moreno-Munoz A, Domingo-Perez F, et al. (2012) Smart grid inverter interface: statistical approach applied to event detection. 2012 IEEE International Workshop on Applied Measurements for Power Systems (AMPS) Proceedings, Aachen, Germany. |
| [10] | Abella MA, Chenlo F (2004) Choosing the right inverter for grid-connected PV systems. Renew Energ World 7: 132-147. |
| [11] | Pal D, Bajpai P (2016) Active and reactive power control in three phase solar PV inverter using modified IC method. 2016 21st Century Energy Needs-Materials, Systems and Applications (ICTFCEN), Kharagpur, India. |
| [12] | Jia Y, Wu R (2016) Voltage source grid-connected pv inverters based on mppt and droop control. 2016 IEEE 2nd Annual Southern Power Electronics Conference (SPEC), Auckland, New Zealand. |
| [13] |
Notton G, Lazarov V, Stoyanov L (2010) Optimal sizing of a grid-connected PV system for various PV module technologies and inclinations, inverter efficiency characteristics and locations. Renew Energ 35: 541-554. doi: 10.1016/j.renene.2009.07.013
|
| [14] | Dong Y, Dong W, Xu L, et al. (2015) Investigation of PV inverter MPPT efficiency test platform. International Conference on Renewable Power Generation (RPG), 1-4. |
| [15] |
Yazdanpanahi H, Li YW, Xu W (2012) A new control strategy to mitigate the impact of inverter-based DGs on protection system. IEEE T Smart grid 3: 1427-1436. doi: 10.1109/TSG.2012.2184309
|
| [16] | Basso T (2015) IEEE 1547 standards for grid integration of distributed energy resources. IEEE PES Seattle Chapter Meeting, 5-18. |
| [17] | Basso T (2014) IEEE 1547 and 2030 standards for distributed energy resources interconnection and interoperability with the electricity grid. National Renewable Energy Lab, Golden, CO (United States). |
| [18] |
González R, Gubía E, López J, et al. (2008) Transformerless single-phase multilevel-based photovoltaic inverter. IEEE T Ind Electron 55: 2694-2702. doi: 10.1109/TIE.2008.924015
|
| [19] | Dissanayaka D, Fernando CTJ, Abeyratne SG (2011) Smart meter based inverter controlling network for demand response applications in smart grids. 2011 6th International Conference on Industrial and Information Systems, Kandy, Sri Lanka. |
| [20] | Smith JW, Sunderman W, Dugan R, et al. (2011) Smart inverter volt/var control functions for high penetration of PV on distribution systems, Phoenix, USA. |
| [21] | Electric Power Research Institute (2016) Common Functions for Smart Inverters: 4th Edition. Available from: https://www.epri.com/#/pages/product/000000003002008217/. |
| [22] |
Sachin J, Vivek A (2007) A single-stage grid connected inverter topology for solar PV systems with maximum power point tracking. IEEE T Power Electron 22: 1928-1940. doi: 10.1109/TPEL.2007.904202
|
| [23] | Bobruk J (2017) What are "smart" inverters? Presented at Smart Inverters 101, ISEA Business Member Webinar: 5-39. Available from: https://www.illinoissolar.org/resources/Documents/ISEA%20Smart%20Inverter%20Webinar.pptx. |
| [24] | Miñambres-Marcos V, Romero-Cadaval E, Guerrero-Martínez M Á, et al. (2012) Three-phase single stage photovoltaic inverter with active filtering capabilities. IECON 2012-38th Annual Conference on IEEE Industrial Electronics Society, Montreal, Canada. |
| [25] | Kumar N, Saha TK, Dey J (2014) Cascaded two level inverter based grid connected photovoltaic system: Modelling and control. 2014 IEEE International Conference on Industrial Technology (ICIT), Busan, South Korea. |
| [26] | Schonardie MF, Ruseler A, Coelho RF, et al. (2010) Three-phase grid-connected PV system with active and reactive power control using dq0 transformation. 2010 9th IEEE/IAS International Conference on Industry Applications-INDUSCON 2010, Sao Paulo, Brazil. |
| [27] | Jafari M, Olowu TO, Sarwat AI (2018) Optimal smart inverters volt-var curve selection with a multi-objective volt-var optimization using evolutionary algorithm approach. 2018 North American Power Symposium, Fargo, USA. |
| [28] | Wang Z, Ren L, Gong C, et al. (2015) The control strategy of interleaved full-bridge inverter with a common DC-bus. 2015 IEEE 10th Conference on Industrial Electronics and Applications. Auckland, New Zealand. |
| [29] | Maknouninejad A, Kutkut N, Batarseh I, et al. (2011) Analysis and control of PV inverters operating in VAR mode at night, Anaheim, USA. |
| [30] | Chakraborty S, Hoke A, Lundstrom B (2015) Evaluation of multiple inverter volt-var control interactions with realistic grid impedances. 2015 IEEE Power and Energy Society General Meeting, Denver, USA. |
| [31] | Braslavsky JH, Collins LD, Ward JK (2017) Voltage stability in a grid-connected inverter with automatic Volt-Watt and Volt-VAR functions. IEEE T Smart Grid 10: 84-94. |
| [32] | Ding F, Nguyen A, Walinga S, et al. (2017) Application of autonomous smart inverter volt-var function for voltage reduction energy savings and power quality in electric distribution systems. 2017 IEEE Power and Energy Society Innovative Smart Grid Technologies Conference, Washington, USA. |
| [33] | Kraiczy M, York B, Bello M, et al. (2018) Coordinating smart inverters with advanced distribution voltage control strategies. 2018 IEEE Power and Energy Society General Meeting, Portland, USA. |
| [34] | Kashani MG, Mobarrez M, Bhattacharya S (2018) Smart inverter volt-watt control design in high PV-penetrated distribution systems. IEEE T Ind Appl 55: 1147-1156. |
| [35] | Kerekes T, Teodorescu R, Rodríguez P, et al. (2009) A new high-efficiency single-phase transformerless PV inverter topology. IEEE T Ind Electron 58: 184-191. |
| [36] | Yang T, Mok KT, Tan SC, et al. (2016) Electric springs with coordinated battery management for reducing voltage and frequency fluctuations in microgrids. IEEE T Smart Grid 9: 1943-1952. |
| [37] | Brian S, Aminul H, Lindsey R, et al. (2016) Smart grid ready PV inverters with utility communication. Electric Power Research Institute 1-56. |
| [38] | Liang Z, Alesi L, Zhou X, et al. (2010) Digital controller development for grid-tied photovoltaic inverter with model based technique. 2010 Twenty-Fifth Annual IEEE Applied Power Electronics Conference and Exposition, Palm Springs, USA. |
| [39] |
Qi J, Hahn A, Lu X, et al. (2016) Cybersecurity for distributed energy resources and smart inverters. IET Cyber-Physical Syst Theory Appl 1: 28-39. doi: 10.1049/iet-cps.2016.0018
|
| [40] | Ortjohann E, Lingemann M, Mohd A, et al. (2018) A general architecture for modular smart inverters. 2008 IEEE International Symposium on Industrial Electronics, Cambridge, UK. |
| [41] | Xue Y, Chang L, Song P (2004) Recent developments in topologies of single-phase buck-boost inverters for small distributed power generators: an overview. The 4th International Power Electronics and Motion Control Conference, Xi'an, China. |
| [42] | Xue Y, Guerrero JM (2015) Smart inverters for utility and industry applications. Proceedings of PCIM Europe 2015; International Exhibition and Conference for Power Electronics, Intelligent Motion, Renewable Energy and Energy Management, Nuremberg, Germany. |
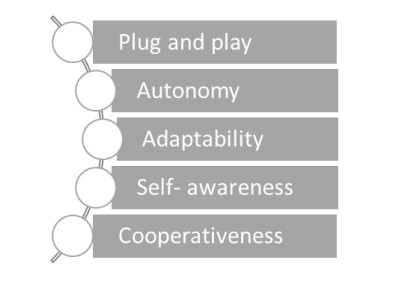
| [43] | Zeng B, Kong XY, Zhang YS, et al. (2015) Study on plug and play access control for distributed generation in microgrid. 2015 International Conference on Electrical, Automation and Mechanical Engineering, Atlantis Press. |
| [44] | Mitchell M (2005) Self-awareness and control in decentralized systems. AAAI Spring Symposium: Metacognition in Computation, Portland, USA. |
| [45] |
Arbab-Zavar B, Palacios-Garcia EJ, Vasquez JC, et al. (2019) Smart inverters for microgrid applications: a review. Energies 12: 840. doi: 10.3390/en12050840
|
| [46] |
Nutkani IU, Loh PC, Wang P, et al. (2014) Autonomous droop scheme with reduced generation cost. IEEE T Ind Electron 61: 6803-6811. doi: 10.1109/TIE.2014.2314056
|
| [47] | CPUC Smart Inverter Working Group (2014) Recommendations for updating the technical requirements for inverters in distributed energy resources: 5-72. Available from: https://www.cpuc.ca.gov/WorkArea/DownloadAsset.aspx?id=3189. |
| [48] |
Yang Y, Blaabjerg F, Wang H, et al. (2016) Power control flexibilities for grid-connected multi-functional photovoltaic inverters. IET Renew Power Gen 10: 504-513. doi: 10.1049/iet-rpg.2015.0133
|
| [49] | Bouzguenda M, Gastli A, Al Badi AH, et al. (2011) Solar photovoltaic inverter requirements for smart grid applications. 2011 IEEE PES Conference on Innovative Smart Grid Technologies-Middle East, Jeddah, Saudi Arabia. |
| [50] |
Liang H, Choi BJ, Zhuang W, et al. (2013) Stability enhancement of decentralized inverter control through wireless communications in microgrids. IEEE T Smart Grid 4: 321-331. doi: 10.1109/TSG.2012.2226064
|
| [51] | Ishii H, Hayashi Y (2017) Optimal smart functions of large-scale PV inverters in distribution systems. 2017 IEEE Innovative Smart Grid Technologies-Asia, Auckland, New Zealand. |
| [52] | Zhao X, Chang L, Shao R, et al. (2018) Power system support functions provided by smart inverters-A review. CPSS T Power Electron Appl 3: 25-35. |
| [53] | Kortenbruck J, Ortjohann E, Leksawat S, et al. (2014) Advanced control functions for smart grid inverter under asymmetrical conditions. 3rd Renewable Power Generation Conference, Naples, Italy. |

Figures(8) / Tables(7)

Kushan Tharuka Lulbadda, K. T. M. U. Hemapala. The additional functions of smart inverters[J]. AIMS Energy, 2019, 7(6): 971-988. doi: 10.3934/energy.2019.6.971







 DownLoad:
DownLoad: