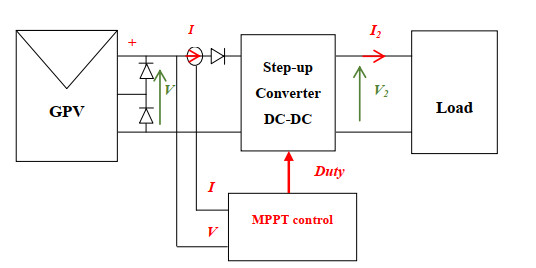
A novel approach defined by the artificial neural network (ANN) model trained by the improved Gauss-Newton in conjunction with a simulated annealing technique is used to control a step-up converter. To elucidate the superiority of this innovative method and to show its high precision and speed in achieving the right value of the Maximum Power Point (MPP), a set of three comparative Maximum Power Point Tracker (MPPT) methods (Perturbation and observation, ANN and ANN associated with perturbation and observation) are exanimated judiciously. The behavior of these methods is observed and tested for a fixed temperature and irradiance. As a result, the proposed approach quickly tracks the right MPP = 18.59 W in just 0.04382 s. On the other hand, the outstanding ability of the suggested method is demonstrated by varying the irradiance values (200 W/m2, 300 W/m2, 700 W/m2, 1000 W/m2, 800 W/m2 and 400 W/m2) and by varying the temperature values (15℃, 35℃, 45℃ and 5℃). Therefore, the ANN trained by Gauss-Newton in conjunction with simulated annealing shows a high robustness and achieves the correct value of MPP for each value of irradiance with an efficiency 99.54% and for each value of temperature with an efficiency 99.98%; the three other methods sometimes struggle to achieve the right MPP for certain irradiance values and often remains stuck in its surroundings.
Citation: Fayrouz Dkhichi. Improved MPPT algorithm: Artificial neural network trained by an enhanced Gauss-Newton method[J]. AIMS Electronics and Electrical Engineering, 2023, 7(4): 380-405. doi: 10.3934/electreng.2023020
A novel approach defined by the artificial neural network (ANN) model trained by the improved Gauss-Newton in conjunction with a simulated annealing technique is used to control a step-up converter. To elucidate the superiority of this innovative method and to show its high precision and speed in achieving the right value of the Maximum Power Point (MPP), a set of three comparative Maximum Power Point Tracker (MPPT) methods (Perturbation and observation, ANN and ANN associated with perturbation and observation) are exanimated judiciously. The behavior of these methods is observed and tested for a fixed temperature and irradiance. As a result, the proposed approach quickly tracks the right MPP = 18.59 W in just 0.04382 s. On the other hand, the outstanding ability of the suggested method is demonstrated by varying the irradiance values (200 W/m2, 300 W/m2, 700 W/m2, 1000 W/m2, 800 W/m2 and 400 W/m2) and by varying the temperature values (15℃, 35℃, 45℃ and 5℃). Therefore, the ANN trained by Gauss-Newton in conjunction with simulated annealing shows a high robustness and achieves the correct value of MPP for each value of irradiance with an efficiency 99.54% and for each value of temperature with an efficiency 99.98%; the three other methods sometimes struggle to achieve the right MPP for certain irradiance values and often remains stuck in its surroundings.

| [1] |
Saccardo RR, Domingues AM, Battistelle RAG, Bezerra BS, Siqueira RM, dos Santos Neto JB (2023) Investment in photovoltaic energy: An attempt to frame Brazil within the 2030 passage target of the Paris agreement. Cleaner Energy Systems 5: 100070. https://doi.org/10.1016/j.cles.2023.100070 doi: 10.1016/j.cles.2023.100070

|
| [2] |
Ma Z, Hu L, Mao H, Shao Q, Tian Z, Luo Y, et al. (2023) Shading effect and energy-saving potential of rooftop photovoltaic on the top-floor room. Solar Energy 265: 112099. https://doi.org/10.1016/j.solener.2023.112099 doi: 10.1016/j.solener.2023.112099
|
| [3] |
Junior RNY, Ochoa AAV, Leite GDNP, Silva HCN, da Costa JA, Tiba C, et al. (2023) Real-time energy and economic performance of the multi-zone photovoltaic-drive air conditioning system for an office building in a tropical climate. Energ Convers Manage 297: 117713. https://doi.org/10.1016/j.enconman.2023.117713 doi: 10.1016/j.enconman.2023.117713
|
| [4] |
Yin Y, Liu J (2023) Collaborative decision-making model for capacity allocation of photovoltaics energy storage system under Energy Internet in China. J Energy Storage 66: 107456. https://doi.org/10.1016/j.est.2023.107456 doi: 10.1016/j.est.2023.107456
|
| [5] |
Traiki G, El Magri A, Lajouad R, Bouattane O (2023) Multi-objective control and optimization of a stand-alone photovoltaic power conversion system with battery storage energy management. IFAC Journal of Systems and Control 26: 100227. https://doi.org/10.1016/j.ifacsc.2023.100227 doi: 10.1016/j.ifacsc.2023.100227
|
| [6] |
Burhan M, Chua KJE, Ng KC (2016) Sunlight to hydrogen conversion: Design optimization and energy management of concentrated photovoltaic (CPV-Hydrogen) system using micro genetic algorithm. Energy 99: 115-128. https://doi.org/10.1016/j.energy.2016.01.048 doi: 10.1016/j.energy.2016.01.048
|
| [7] |
Das D, Panda DP, Tongbram B, Saha J, Chavan V, Chakrabarti S(2018) Optimization of hybrid InAs stranski krastanov and submonolayer quantum dot heterostructures and its effect on photovoltaic energy conversion efficiency in near infrared region. Solar Energy 171: 64-72. https://doi.org/10.1016/j.solener.2018.06.030 doi: 10.1016/j.solener.2018.06.030
|
| [8] |
Corrado C, Leow SW, Osborn M, Chan E, Balaban B, Carter SA (2013) Optimization of gain and energy conversion efficiency using front-facing photovoltaic cell luminescent solar concentrator design. Solar Energy Materials and Solar Cells 111: 74-81. https://doi.org/10.1016/j.solmat.2012.12.030 doi: 10.1016/j.solmat.2012.12.030
|
| [9] |
Riyadi TWB, Effendy M, Utomo BR, Wijayanta AT (2023) Performance of a photovoltaic-thermoelectric generator panel in combination with various solar tracking systems. Appl Therm Eng 253: 121336. https://doi.org/10.1016/j.applthermaleng.2023.121336 doi: 10.1016/j.applthermaleng.2023.121336
|
| [10] |
Baghaz E, Melhaoui M, Yaden F, Hirech K, Kassmi K (2013) Design realization and optimization of the photovoltaic systems equipped with analog and digital MPPT commands. Energy Procedia 42: 270-279. https://doi.org/10.1016/j.egypro.2013.11.027 doi: 10.1016/j.egypro.2013.11.027
|
| [11] |
Masmoudi A, Abdelkafi A, Krichen L, Saidi AS (2022) An experimental approach for improving stability in DC bus voltage of a stand-alone photovoltaic generator. Energy 257: 124797. https://doi.org/10.1016/j.energy.2022.124797 doi: 10.1016/j.energy.2022.124797
|
| [12] |
Choi WY, Lee CG (2012) Photovoltaic panel integrated power conditioning system using a high efficiency step-up DC–DC converter. Renewable Energy 41: 227-234. https://doi.org/10.1016/j.renene.2011.10.023 doi: 10.1016/j.renene.2011.10.023
|
| [13] |
Nakpin A, Khwan S (2016) A Novel High Step-up DC-DC Converter for Photovoltaic Applications. Procedia Computer Science 86: 409-412. https://doi.org/10.1016/j.procs.2016.05.051 doi: 10.1016/j.procs.2016.05.051
|
| [14] |
Cha WJ, Kwon JM, Kwon BH (2016) Highly efficient step-up dc–dc converter for photovoltaic micro-inverter. Solar Energy 135: 14-21. https://doi.org/10.1016/j.solener.2016.05.024 doi: 10.1016/j.solener.2016.05.024
|
| [15] |
Mustafa Y, Yin H, Lu Y, Ruderman A (2021) Constrained minimization of switched capacitor converter equivalent resistance by adjusting transistor sizes and duty cycles. Microelectronics Journal 112: 105061. https://doi.org/10.1016/j.mejo.2021.105061 doi: 10.1016/j.mejo.2021.105061
|
| [16] |
Hafiz M, Ahmed KZ, Islam D, Rashid ABMH (2011) Design and implementation of a 0.8 V input, 84% duty cycle, variable frequency step-up converter. Microelectron J 42: 648-660. https://doi.org/10.1016/j.mejo.2011.03.005 doi: 10.1016/j.mejo.2011.03.005
|
| [17] |
Amir A, Amir A, Seng CH, el Khateb A, Selvaraj J, Rahim NA (2018) Application of modified classical numerical methods for DMPPT on Buck and Boost converters. Solar Energy 173: 437-448. https://doi.org/10.1016/j.solener.2018.07.088 doi: 10.1016/j.solener.2018.07.088
|
| [18] |
Baba AO, Liu G, Chen X (2020) Classification and Evaluation Review of Maximum Power Point Tracking Methods. Sustainable Futures 2: 100020. https://doi.org/10.1016/j.sftr.2020.100020 doi: 10.1016/j.sftr.2020.100020
|
| [19] |
Pandey NK, Pachauri RK, Choudhury S, Sahu RK (2023) Asymmetrical interval Type-2 Fuzzy logic controller based MPPT for PV system under sudden irradiance changes. Materials today proceeding 80: 710-716. https://doi.org/10.1016/j.matpr.2022.11.074 doi: 10.1016/j.matpr.2022.11.074
|
| [20] |
Ullah K, Ishaq M, Tchier F, Ahmad H, Ahmad Z (2023) Fuzzy-based maximum power point tracking (MPPT) control system for photovoltaic power generation system. Results in Engineering 20: 101466. https://doi.org/10.1016/j.rineng.2023.101466 doi: 10.1016/j.rineng.2023.101466
|
| [21] |
Mohammadinodoushan M, Abbassi R, Jerbi H, Ahmed FW, Rezvani A (2021) A new MPPT design using variable step size perturb and observe method for PV system under partially shaded conditions by modified shuffled frog leaping algorithm- SMC controller. Sustain Energy Techn 45: 101056. https://doi.org/10.1016/j.seta.2021.101056 doi: 10.1016/j.seta.2021.101056
|
| [22] |
Loukriz A, Haddadi M, Messalt S (2016) Simulation and experimental design of a new advanced variable step size Incremental Conductance MPPT algorithm for PV system. ISA Transactions 62: 30-38. https://doi.org/10.1016/j.isatra.2015.08.006 doi: 10.1016/j.isatra.2015.08.006
|
| [23] |
Goel A, Goel AK, Kumar A (2022) The role of artificial neural network and machine learning in utilizing spatial information. Spat Inf Res 31: 275-285. https://doi.org/10.1007/s41324-022-00494-x doi: 10.1007/s41324-022-00494-x
|
| [24] |
Toumi D, Attous DB, Ibrahim A, Tarek B (2022) Maximum power point tracking of photovoltaic array using fuzzy logic control. International Journal of Power Electronics and Drive Systems 13: 2440. https://doi.org/10.11591/ijpeds.v13.i4.pp2440-2449 doi: 10.11591/ijpeds.v13.i4.pp2440-2449
|
| [25] | Manna S, Singh DK, Akella AK (2023) Hybrid two-stage adaptive maximum power point tracking for stand-alone, grid integration, and partial shaded PV system. Int J Adapt Control. https://doi.org/10.1002/acs.3684 |
| [26] |
Manna S, Akella AK, Singh DK (2023) Novel Lyapunov-based rapid and ripple-free MPPT using a robust model reference adaptive controller for solar PV system. Prot Contr Mod Pow Syst 8: 1-25. https://doi.org/10.1186/s41601-023-00288-9 doi: 10.1186/s41601-023-00288-9
|
| [27] |
Singh DK, Akella AK, Manna S (2023) Adjustable variable step-based MRAC MPPT for solar PV system in highly fluctuating and cloudy atmospheric conditions. Electrical Engineering 105: 3751-3772. https://doi.org/10.1007/s00202-023-01922-3 doi: 10.1007/s00202-023-01922-3
|
| [28] |
Ahmed R, Namaane A, M'Sirdi NK (2013) Improvement in Perturb and Observe Method Using State Flow Approach. Energy Procedia 42: 614-623. https://doi.org/10.1016/j.egypro.2013.11.063 doi: 10.1016/j.egypro.2013.11.063
|
| [29] |
Argyros IK, Magreñ án AA (2014) Local convergence analysis of proximal Gauss–Newton method for penalized nonlinear least squares problems. Appl Math Comput 241: 401-408. https://doi.org/10.1016/j.amc.2014.04.087 doi: 10.1016/j.amc.2014.04.087
|
| [30] |
Dkhichi F, Oukarfi B, Fakkar A, Belbounaguia N (2014) Parameter identification of solar cell model using Levenberg–Marquardt algorithm combined with simulated annealing. Solar Energy 110: 781-788. https://doi.org/10.1016/j.solener.2014.09.033 doi: 10.1016/j.solener.2014.09.033
|
| [31] |
Bao T, Li Z, Pu O, Chan RWK, Zhao Z, Pan Y, et al. (2023) Modal analysis of tracking photovoltaic support system. Solar Energy 265: 112088. https://doi.org/10.1016/j.solener.2023.112088 doi: 10.1016/j.solener.2023.112088
|
| [32] |
Ouedraogo A, Diallo A, Goro S, Ilboudo WDA, Madougou S, Bathiebo DJ, et al. (2022) Analysis of the solar power plant efficiency installed in the premises of a hospital — Case of the Pediatric Charles De Gaulle of Ouagadougou. Solar Energy 241: 120-129. https://doi.org/10.1016/j.solener.2022.05.051 doi: 10.1016/j.solener.2022.05.051
|
| [33] |
Muhammadsharif FF, Hashim S, Hameed SS, Ghoshal SK, Abdullah IK, Macdonald JE, et al. (2019) Brent's algorithm based new computational approach for accurate determination of single-diode model parameters to simulate solar cells and modules. Solar Energy 193: 782-798. https://doi.org/10.1016/j.solener.2019.09.096 doi: 10.1016/j.solener.2019.09.096
|
| [34] |
Ndi FE, Perabi SN, Ndjakomo SE, Abessolo GO, Mengata GM(2021) Estimation of single-diode and two diode solar cell parameters by equilibrium optimizer method. Energy Reports 7: 4761-4768. https://doi.org/10.1016/j.egyr.2021.07.025 doi: 10.1016/j.egyr.2021.07.025
|
| [35] |
Zhang SMF, Seif JP, Abbott MD, Le AH, Allen TG, Perez-Wurfl I, et al. (2022) Illumination-dependent temperature coefficients of the electrical parameters of modern silicon solar cell architectures. Nano Energy 98: 107221. https://doi.org/10.1016/j.nanoen.2022.107221 doi: 10.1016/j.nanoen.2022.107221
|
| [36] |
Park C, Balaji N, Ahn S, Park J, Cho E, Yi J (2020) Effects of tunneling oxide defect density and inter-diffused carrier concentration on carrier selective contact solar cell performance: Illumination and temperature effects. Solar Energy 211: 62-73. https://doi.org/10.1016/j.solener.2020.09.060 doi: 10.1016/j.solener.2020.09.060
|
| [37] | Luo P, Pan J, Hong J, Liang J, Jiang H, Yang D (2023) An ultrahigh synchronous drive step-up converter for PEMFC and its explicit model predictive control: A neural network fitting strategy. Int J Hydrogen Energy. https://doi.org/10.1016/j.ijhydene.2023.08.113 |
| [38] |
Li P, Wang Y, Zuo Z (2023) Robust multiple frequency design on voltage-mode control of DC-DC boost converters. J Franklin I 360: 1207-1225. https://doi.org/10.1016/j.jfranklin.2022.12.017 doi: 10.1016/j.jfranklin.2022.12.017
|
| [39] | Morales DS (2010) Maximum Power Point Tracking Algorithms for Photovoltaic Applications, Faculty of Electronics, Communications and Automation. Master of Science in Technology. |
| [40] |
Naidu IES, Srikanth S, Rao A, Venkatanarayana A (2023) A novel mine blast optimization algorithm (MBOA) based MPPT controlling for grid-PV systems. AIMS Electronics and Electrical Engineering 7: 135-155. https://doi.org/10.3934/electreng.2023008 doi: 10.3934/electreng.2023008
|
| [41] |
Sivakumar P, Kader AA, Kaliavaradhan Y, Arutchelvi M (2015) Analysis and enhancement of PV efficiency with incremental conductance MPPT technique under non-linear loading conditions. Renew Energ 81: 543-550. https://doi.org/10.1016/j.renene.2015.03.062 doi: 10.1016/j.renene.2015.03.062
|
| [42] |
Wu Y, Jakobsson A, Liu L (2023) Super-resolution Direction of Arrival Estimation Using a Minimum Mean-Square Error Framework. Signal Processing 212: 109164. https://doi.org/10.1016/j.sigpro.2023.109164 doi: 10.1016/j.sigpro.2023.109164
|
| [43] | Dkhichi F, Oukarfi B, El Kouari Y, Ouoba D, Fakkar A, Sabiri Z (2016) Performances of Artificial Neural Network combined with Perturb & Observe technique in maximizing the photovoltaic system power. International Renewable and Sustainable Energy Conference (IRSEC), 951-955. https://doi.org/10.1109/IRSEC.2016.7984023 |
| [44] |
Kassem AM (2012) MPPT control design and performance improvements of a PV generator powered DC motor-pump system based on artificial neural networks. Electrical Power and Energy Systems 43: 90–98. https://doi.org/10.1016/j.ijepes.2012.04.047 doi: 10.1016/j.ijepes.2012.04.047
|
| [45] |
Ansari QH, Uddin M, Yao JC (2024) Convergence of the Gauss-Newton method for convex composite optimization problems under majorant condition on Riemannian manifolds. J Complexity 80: 101788. https://doi.org/10.1016/j.jco.2023.101788 doi: 10.1016/j.jco.2023.101788
|
| [46] | Cyril Lahore (2012) Optimisation de commandes MPPT. Available from : https://dumas.ccsd.cnrs.fr/dumas-01304277. |























Figures(24) / Tables(4)

Fayrouz Dkhichi. Improved MPPT algorithm: Artificial neural network trained by an enhanced Gauss-Newton method[J]. AIMS Electronics and Electrical Engineering, 2023, 7(4): 380-405. doi: 10.3934/electreng.2023020







 DownLoad:
DownLoad: