Citation: Moazameh Golshani, Mohsen Basiri, Mohammad Shabani, Iraj Aghaei, Majid Asadi-Shekaari. Effects of erythropoietin on bile duct ligation-induced neuro-inflammation in male rats[J]. AIMS Neuroscience, 2019, 6(2): 43-53. doi: 10.3934/Neuroscience.2019.2.43

| [1] |
Felipo V (2013) Hepatic encephalopathy: effects of liver failure on brain function. Nat Rev Neurosci 14: 851–858. doi: 10.1038/nrn3587

|
| [2] |
Olde Damink SW, Jalan R, Dejong CH (2009) Interorgan ammonia trafficking in liver disease. Metab Brain Dis 24: 169–181. doi: 10.1007/s11011-008-9122-5
|
| [3] |
Butterworth RF, Norenberg MD, Felipo V, et al. (2009) Experimental models of hepatic encephalopathy: ISHEN guidelines. Liver Int 29: 783–788. doi: 10.1111/j.1478-3231.2009.02034.x
|
| [4] |
Leke R, Oliveira DL, Forgiarini LF, et al. (2013) Impairment of short term memory in rats with hepatic encephalopathy due to bile duct ligation. Metab Brain Dis 28: 187–192. doi: 10.1007/s11011-012-9347-1
|
| [5] |
Weissenborn K, Giewekemeyer K, Heidenreich S, et al. (2005) Attention, memory, and cognitive function in hepatic encephalopathy. Metab Brain Dis 20: 359–367. doi: 10.1007/s11011-005-7919-z
|
| [6] |
Ferenci P, Lockwood A, Mullen K, et al. (2002) Hepatic encephalopathy--definition, nomenclature, diagnosis, and quantification: final report of the working party at the 11th World Congresses of Gastroenterology, Vienna, 1998. Hepatology 35: 716–721. doi: 10.1053/jhep.2002.31250
|
| [7] | Patel A, Wade JB, Thacker LR, et al. (2014) 991 Brain Reserve Modulates Health-Related Quality of Life in Patients With Cirrhosis Independent of Covert Hepatic Encephalopathy and MELD Score. Gastroenterology 146: S–932. |
| [8] |
Rose CF (2012) Ammonia-lowering strategies for the treatment of hepatic encephalopathy. Clin Pharmacol Ther 92: 321–331. doi: 10.1038/clpt.2012.112
|
| [9] |
Savlan I, Liakina V, Valantinas J (2014) Concise review of current concepts on nomenclature and pathophysiology of hepatic encephalopathy. Medicina (Kaunas) 50: 75–81. doi: 10.1016/j.medici.2014.06.008
|
| [10] |
Rama Rao KV, Jayakumar AR, Norenberg MD (2012) Glutamine in the pathogenesis of acute hepatic encephalopathy. Neurochem Int 61: 575–580. doi: 10.1016/j.neuint.2012.01.012
|
| [11] |
Rothman DL, De Feyter HM, Maciejewski PK, et al. (2012) Is there in vivo evidence for amino acid shuttles carrying ammonia from neurons to astrocytes? Neurochem Res 37: 2597–2612. doi: 10.1007/s11064-012-0898-7
|
| [12] |
Jover R, Rodrigo R, Felipo V, et al. (2006) Brain edema and inflammatory activation in bile duct ligated rats with diet-induced hyperammonemia: A model of hepatic encephalopathy in cirrhosis. Hepatology 43: 1257–1266. doi: 10.1002/hep.21180
|
| [13] |
Sheen JM, Huang LT, Hsieh CS, et al. (2010) Bile duct ligation in developing rats: temporal progression of liver, kidney, and brain damage. J Pediatr Surg 45: 1650–1658. doi: 10.1016/j.jpedsurg.2009.12.019
|
| [14] | Aldridge DR, Tranah EJ, Shawcross DL (2015) Pathogenesis of hepatic encephalopathy: role of ammonia and systemic inflammation. J Clin Exp Hepatol 5: S7–S20. |
| [15] |
Rodrigo R, Cauli O, Gomez-Pinedo U, et al. (2010) Hyperammonemia induces neuroinflammation that contributes to cognitive impairment in rats with hepatic encephalopathy. Gastroenterology 139: 675–684. doi: 10.1053/j.gastro.2010.03.040
|
| [16] | Jayakumar AR, Rama Rao KV, Norenberg MD (2015) Neuroinflammation in hepatic encephalopathy: mechanistic aspects. J Clin Exp Hepatol 5: S21–28. |
| [17] |
Butterworth RF (2012) Reprint of: Neuroinflammation in acute liver failure: Mechanisms and novel therapeutic targets. Neurochem Int 60: 715–722. doi: 10.1016/j.neuint.2012.03.014
|
| [18] |
Aghaei I, Nazeri M, Shabani M, et al. (2015) Erythropoietin ameliorates the motor and cognitive function impairments in a rat model of hepatic cirrhosis. Metab Brain Dis 30: 197–204. doi: 10.1007/s11011-014-9600-x
|
| [19] |
Aghaei I, Shabani M, Doustar N, et al. (2014) Peroxisome proliferator-activated receptor-gamma activation attenuates motor and cognition impairments induced by bile duct ligation in a rat model of hepatic cirrhosis. Pharmacol Biochem Behav 120: 133–139. doi: 10.1016/j.pbb.2014.03.002
|
| [20] |
Nairz M, Schroll A, Moschen AR, et al. (2011) Erythropoietin contrastingly affects bacterial infection and experimental colitis by inhibiting nuclear factor-kappaB-inducible immune pathways. Immunity 34: 61–74. doi: 10.1016/j.immuni.2011.01.002
|
| [21] | Bond WS, Rex TS (2014) Evidence that erythropoietin modulates neuroinflammation through differential action on neurons, astrocytes, and microglia. Front Immunol 5: 523. |
| [22] |
Aghaei I, Hajali V, Dehpour A, et al. (2016) Alterations in the intrinsic electrophysiological properties of Purkinje neurons in a rat model of hepatic encephalopathy: Relative preventing effect of PPARgamma agonist. Brain Res Bull 121: 16–25. doi: 10.1016/j.brainresbull.2015.12.002
|
| [23] |
Shabani M, Ebrahimpoor F, Firouzjaei MA, et al. (2019) Modulation of sphingosine-1-phosphate receptor by FTY720 contributes in improvement of hepatic encephalopathy induced by bile duct ligation. Brain Res Bull 146: 253–269. doi: 10.1016/j.brainresbull.2019.01.012
|
| [24] |
Tahamtan M, Aghaei I, Pooladvand V, et al. (2017) Characterization of the CA1 pyramidal neurons in rat model of hepatic cirrhosis: insights into their electrophysiological properties. Metab Brain Dis 32: 881–889. doi: 10.1007/s11011-017-9966-7
|
| [25] |
Onoda A, Takeda K, Umezawa M (2017) Dose-dependent induction of astrocyte activation and reactive astrogliosis in mouse brain following maternal exposure to carbon black nanoparticle. Part Fibre Toxicol 14: 4. doi: 10.1186/s12989-017-0184-6
|
| [26] |
Matsui H, Ohgomori T, Natori T, et al. (2013) Keratan sulfate expression in microglia is diminished in the spinal cord in experimental autoimmune neuritis. Cell Death Dis 4: e946. doi: 10.1038/cddis.2013.479
|
| [27] |
Javadi-Paydar M, Ghiassy B, Ebadian S, et al. (2013) Nitric oxide mediates the beneficial effect of chronic naltrexone on cholestasis-induced memory impairment in male rats. Behav Pharmacol 24: 195–206. doi: 10.1097/FBP.0b013e3283618a8c
|
| [28] |
Nasehi M, Piri M, Abbolhasani K, et al. (2013) Involvement of opioidergic and nitrergic systems in memory acquisition and exploratory behaviors in cholestatic mice. Behav Pharmacol 24: 180–194. doi: 10.1097/FBP.0b013e3283618aab
|
| [29] |
Butterworth RF (2011) Hepatic encephalopathy: a central neuroinflammatory disorder? Hepatology 53: 1372–1376. doi: 10.1002/hep.24228
|
| [30] |
Huang LT, Tiao MM, Tain YL, et al. (2009) Melatonin ameliorates bile duct ligation-induced systemic oxidative stress and spatial memory deficits in developing rats. Pediatr Res 65: 176–180. doi: 10.1203/PDR.0b013e31818d5bc7
|
| [31] |
Brines M, Cerami A (2005) Emerging biological roles for erythropoietin in the nervous system. Nat Rev Neurosci 6: 484–494. doi: 10.1038/nrn1687
|
| [32] |
Levitt DG, Levitt MD (2016) Human serum albumin homeostasis: a new look at the roles of synthesis, catabolism, renal and gastrointestinal excretion, and the clinical value of serum albumin measurements. Int J Gen Med 9: 229–255. doi: 10.2147/IJGM.S102819
|
| [33] |
Caillaud C, Mechta M, Ainge H, et al. (2015) Chronic erythropoietin treatment improves diet-induced glucose intolerance in rats. J Endocrinol 225: 77–88. doi: 10.1530/JOE-15-0010
|
| [34] |
Dhanda S, Sandhir R (2015) Role of dopaminergic and serotonergic neurotransmitters in behavioral alterations observed in rodent model of hepatic encephalopathy. Behav Brain Res 286: 222–235. doi: 10.1016/j.bbr.2015.01.042
|
| [35] |
Su YY, Yang GF, Lu GM, et al. (2015) PET and MR imaging of neuroinflammation in hepatic encephalopathy. Metab Brain Dis 30: 31–45. doi: 10.1007/s11011-014-9633-1
|
| [36] |
Chen JR, Wang BN, Tseng GF, et al. (2014) Morphological changes of cortical pyramidal neurons in hepatic encephalopathy. BMC Neurosci 15: 15. doi: 10.1186/1471-2202-15-15
|
| [37] |
Ponce LL, Navarro JC, Ahmed O, et al. (2013) Erythropoietin neuroprotection with traumatic brain injury. Pathophysiology 20: 31–38. doi: 10.1016/j.pathophys.2012.02.005
|
| [38] |
Wenker SD, Chamorro ME, Vittori DC, et al. (2013) Protective action of erythropoietin on neuronal damage induced by activated microglia. FEBS J 280: 1630–1642. doi: 10.1111/febs.12172
|
| [39] |
McPherson RJ, Juul SE (2008) Recent trends in erythropoietin-mediated neuroprotection. Int J Dev Neurosci 26: 103–111. doi: 10.1016/j.ijdevneu.2007.08.012
|
| [40] |
Mofidi A, Bader A, Pavlica S (2011) The use of erythropoietin and its derivatives to treat spinal cord injury. Mini Rev Med Chem 11: 763–770. doi: 10.2174/138955711796355267
|
| [41] | Hu X, Liou AK, Leak RK, et al. (2014) Neurobiology of microglial action in CNS injuries: receptor-mediated signaling mechanisms and functional roles. Prog Neurobiol 119–120: 60–84. |
| [42] |
Wright G, Swain M, Annane D, et al. (2016) Neuroinflammation in liver disease: sessional talks from ISHEN. Metab Brain Dis 31: 1339–1354. doi: 10.1007/s11011-016-9918-7
|
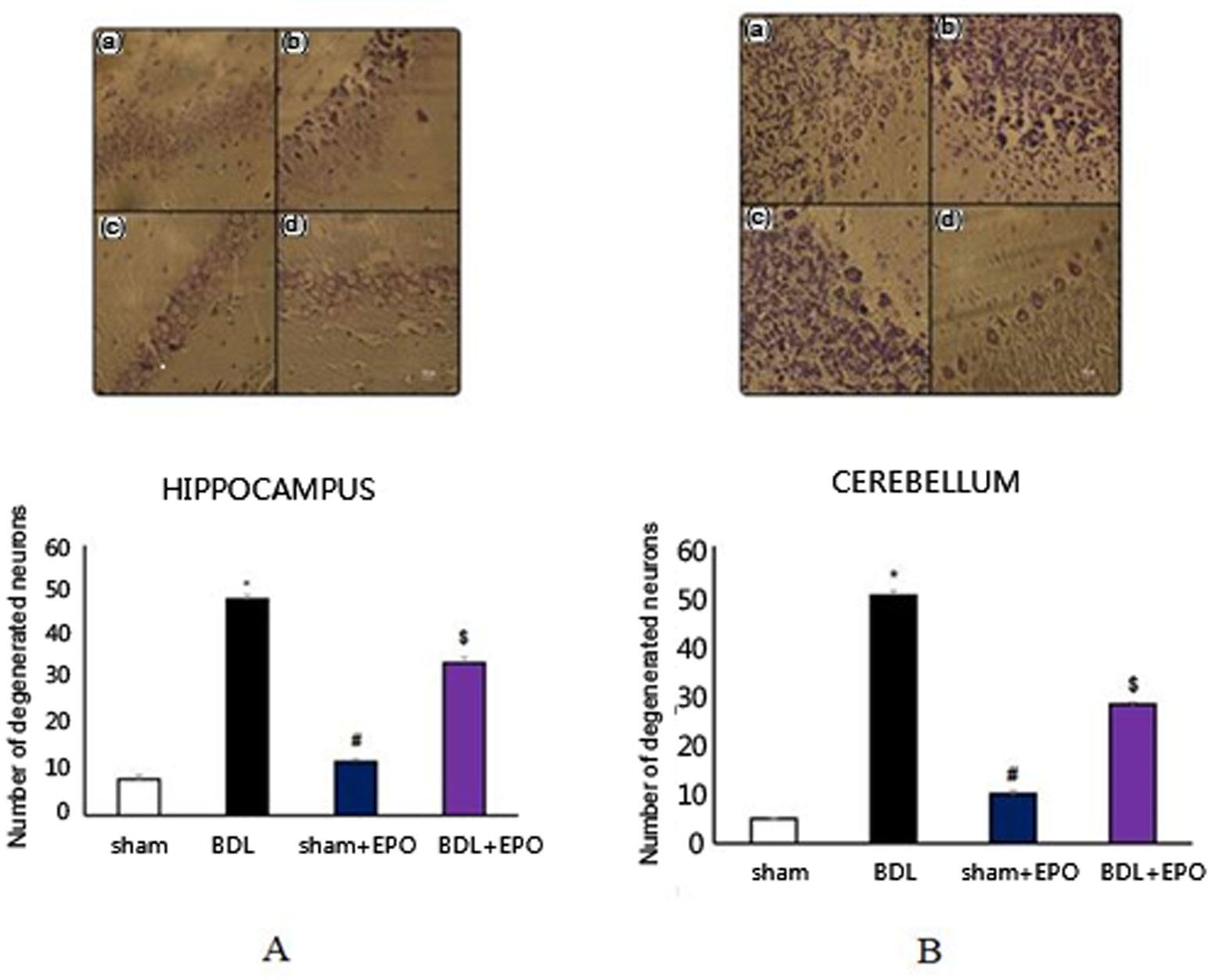
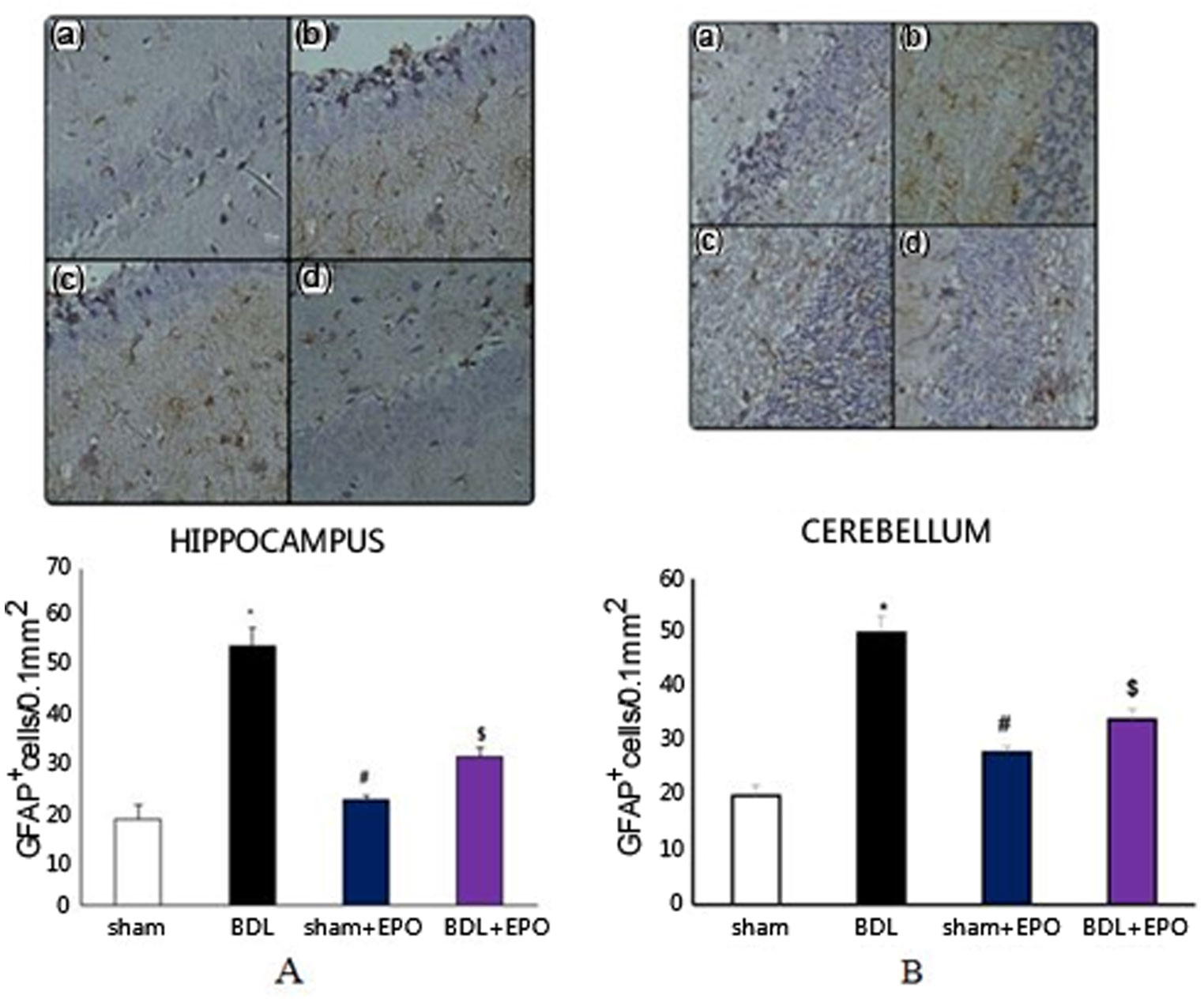
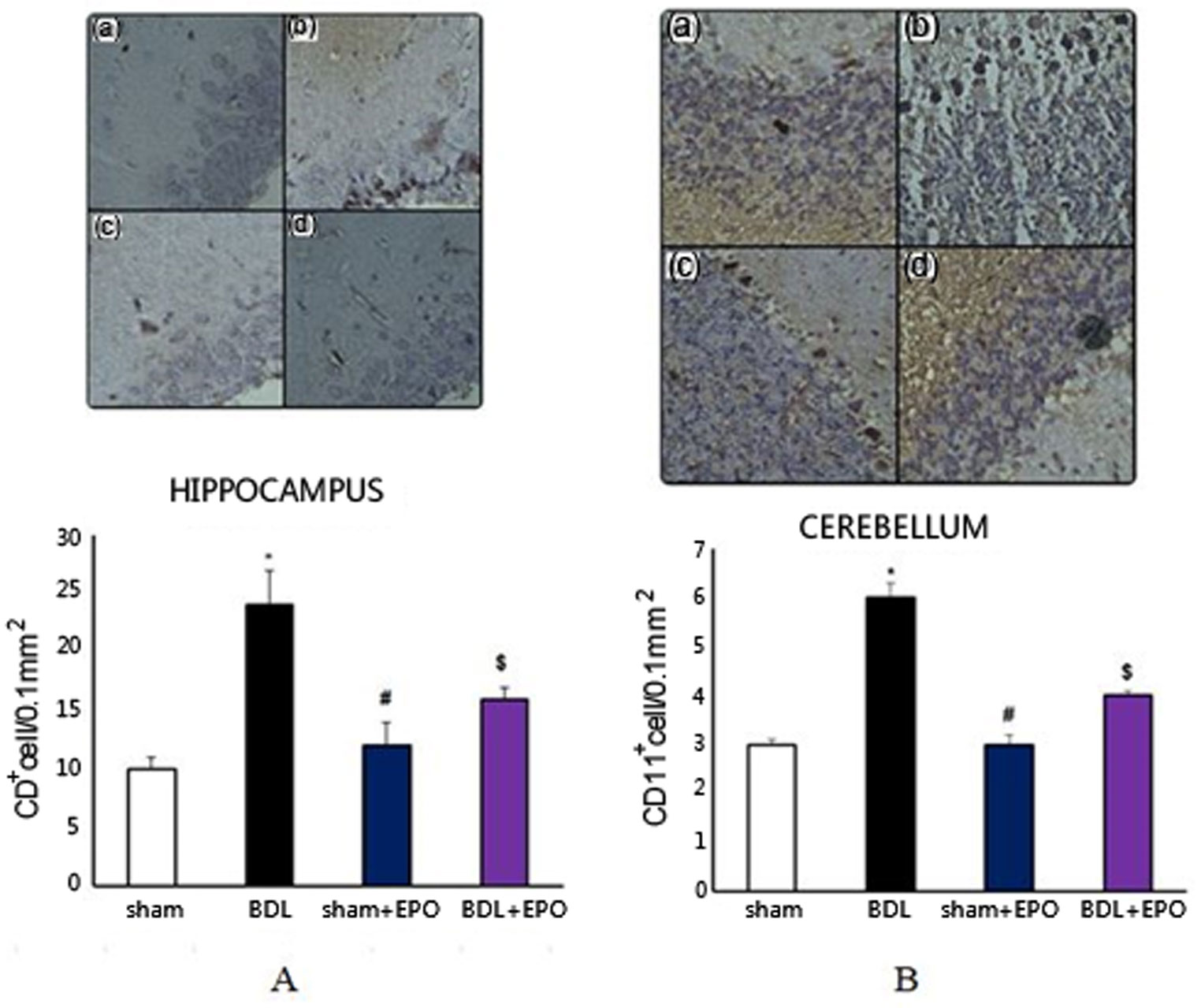
Figures(3) / Tables(1)

Moazameh Golshani, Mohsen Basiri, Mohammad Shabani, Iraj Aghaei, Majid Asadi-Shekaari. Effects of erythropoietin on bile duct ligation-induced neuro-inflammation in male rats[J]. AIMS Neuroscience, 2019, 6(2): 43-53. doi: 10.3934/Neuroscience.2019.2.43







 DownLoad:
DownLoad: