Citation: Karen E. Livermore, Jennifer Munkley, David J. Elliott. Androgen receptor and prostate cancer[J]. AIMS Molecular Science, 2016, 3(2): 280-299. doi: 10.3934/molsci.2016.2.280

| [1] | McNeal JE (1981) The zonal anatomy of the prostate. Prostate 2: 35-49. |
| [2] |
Mcneal JE (1988) Normal Histology of the Prostate. Am J Surg Pathol 12: 619-633. doi: 10.1097/00000478-198808000-00003

|
| [3] | Isaacs JT (1994) Etiology of benign prostatic hyperplasia. Eur Urol 25 Suppl 1: 6-9. |
| [4] | McNeal JE (1978) Origin and evolution of benign prostatic enlargement. Invest Urol 15: 340-345. |
| [5] | Berry SJ, Coffey DS, Walsh PC, et al. (1984) The development of human benign prostatic hyperplasia with age. J Urol 132: 474-479. |
| [6] |
Briganti A, Capitanio U, Suardi N, et al. (2009) Benign Prostatic Hyperplasia and Its Aetiologies. Eur Urol Suppl 8: 865-871. doi: 10.1016/j.eursup.2009.11.002
|
| [7] |
Ferlay J, Soerjomataram I, Dikshit R, et al. (2015) Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer 136: E359-386. doi: 10.1002/ijc.29210
|
| [8] | Cancer Research UK (2013) Prostate cancer statistics. |
| [9] |
Nowell PC (1976) The clonal evolution of tumor cell populations. Science 194: 23-28. doi: 10.1126/science.959840
|
| [10] |
Collins AT, Berry PA, Hyde C, et al. (2005) Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res 65: 10946-10951. doi: 10.1158/0008-5472.CAN-05-2018
|
| [11] |
Visvader JE, Lindeman GJ (2012) Cancer stem cells: current status and evolving complexities. Cell Stem Cell 10: 717-728. doi: 10.1016/j.stem.2012.05.007
|
| [12] |
Ablin RJ, Soanes WA, Bronson P, et al. (1970) Precipitating antigens of the normal human prostate. J Reprod Fertil 22: 573-574. doi: 10.1530/jrf.0.0220573
|
| [13] | Papsidero LD, Wang MC, Valenzuela LA, et al. (1980) A prostate antigen in sera of prostatic cancer patients. Cancer Res 40: 2428-2432. |
| [14] |
Nadler RB, Humphrey PA, Smith DS, et al. (1995) Effect of inflammation and benign prostatic hyperplasia on elevated serum prostate specific antigen levels. J Urol 154: 407-413. doi: 10.1016/S0022-5347(01)67064-2
|
| [15] |
Romero Otero J, Garcia Gomez B, Campos Juanatey F, et al. (2014) Prostate cancer biomarkers: an update. Urol Oncol 32: 252-260. doi: 10.1016/j.urolonc.2013.09.017
|
| [16] |
Gilgunn S, Conroy PJ, Saldova R, et al. (2013) Aberrant PSA glycosylation--a sweet predictor of prostate cancer. Nat Rev Urol 10: 99-107. doi: 10.1038/nrurol.2012.258
|
| [17] |
Hessels D, Klein Gunnewiek JM, van Oort I, et al. (2003) DD3(PCA3)-based molecular urine analysis for the diagnosis of prostate cancer. Eur Urol 44: 8-15. doi: 10.1016/S0302-2838(03)00201-X
|
| [18] |
Attard G, Clark J, Ambroisine L, et al. (2008) Duplication of the fusion of TMPRSS2 to ERG sequences identifies fatal human prostate cancer. Oncogene 27: 253-263. doi: 10.1038/sj.onc.1210640
|
| [19] | Gleason DF, Mellinger GT (1974) Prediction of prognosis for prostatic adenocarcinoma by combined histological grading and clinical staging. J Urol 111: 58-64. |
| [20] | Mellinger GT, Gleason D, Bailar J 3rd (1967) The histology and prognosis of prostatic cancer. J Urol 97: 331-337. |
| [21] |
Conn PM, Crowley WF Jr. (1994) Gonadotropin-releasing hormone and its analogs. Annu Rev Med 45: 391-405. doi: 10.1146/annurev.med.45.1.391
|
| [22] |
Lubahn DB, Joseph DR, Sullivan PM, et al. (1988) Cloning of human androgen receptor complementary DNA and localization to the X chromosome. Science 240: 327-330. doi: 10.1126/science.3353727
|
| [23] |
Gelmann EP (2002) Molecular biology of the androgen receptor. J Clin Oncol 20: 3001-3015. doi: 10.1200/JCO.2002.10.018
|
| [24] |
Edwards A, Hammond HA, Jin L, et al. (1992) Genetic variation at five trimeric and tetrameric tandem repeat loci in four human population groups. Genomics 12: 241-253. doi: 10.1016/0888-7543(92)90371-X
|
| [25] |
Verrijdt G, Haelens A, Claessens F (2003) Selective DNA recognition by the androgen receptor as a mechanism for hormone-specific regulation of gene expression. Mol Genet Metab 78: 175-185. doi: 10.1016/S1096-7192(03)00003-9
|
| [26] |
Maitland NJ, Frame FM, Polson ES, et al. (2011) Prostate cancer stem cells: do they have a basal or luminal phenotype? Horm Cancer 2: 47-61. doi: 10.1007/s12672-010-0058-y
|
| [27] |
Mirosevich J, Bentel JM, Zeps N, et al. (1999) Androgen receptor expression of proliferating basal and luminal cells in adult murine ventral prostate. J Endocrinol 162: 341-350. doi: 10.1677/joe.0.1620341
|
| [28] |
Radmayr C, Lunacek A, Schwentner C, et al. (2008) 5-alpha-reductase and the development of the human prostate. Indian J Urol 24: 309-312. doi: 10.4103/0970-1591.42610
|
| [29] |
Saartok T, Dahlberg E, Gustafsson JA (1984) Relative binding affinity of anabolic-androgenic steroids: comparison of the binding to the androgen receptors in skeletal muscle and in prostate, as well as to sex hormone-binding globulin. Endocrinology 114: 2100-2106. doi: 10.1210/endo-114-6-2100
|
| [30] |
Cano LQ, Lavery DN, Bevan CL (2013) Mini-review: Foldosome regulation of androgen receptor action in prostate cancer. Mol Cell Endocrinol 369: 52-62. doi: 10.1016/j.mce.2013.01.023
|
| [31] |
He B, Kemppainen JA, Voegel JJ, et al. (1999) Activation function 2 in the human androgen receptor ligand binding domain mediates interdomain communication with the NH(2)-terminal domain. J Biol Chem 274: 37219-37225. doi: 10.1074/jbc.274.52.37219
|
| [32] |
Massie CE, Lynch A, Ramos-Montoya A, et al. (2011) The androgen receptor fuels prostate cancer by regulating central metabolism and biosynthesis. EMBO J 30: 2719-2733. doi: 10.1038/emboj.2011.158
|
| [33] |
Cleutjens KB, van Eekelen CC, van der Korput HA, et al. (1996) Two androgen response regions cooperate in steroid hormone regulated activity of the prostate-specific antigen promoter. J Biol Chem 271: 6379-6388. doi: 10.1074/jbc.271.11.6379
|
| [34] |
Sun M, Yang L, Feldman RI, et al. (2003) Activation of phosphatidylinositol 3-kinase/Akt pathway by androgen through interaction of p85alpha, androgen receptor, and Src. J Biol Chem 278: 42992-43000. doi: 10.1074/jbc.M306295200
|
| [35] | Migliaccio A, Castoria G, Di Domenico M, et al. (2000) Steroid-induced androgen receptor-oestradiol receptor beta-Src complex triggers prostate cancer cell proliferation. EMBO J 19: 5406-5417. |
| [36] | Liao RS, Ma S, Miao L, et al. (2013) Androgen receptor-mediated non-genomic regulation of prostate cancer cell proliferation. Transl Androl Urol 2: 187-196. |
| [37] |
Peterziel H, Mink S, Schonert A, et al. (1999) Rapid signalling by androgen receptor in prostate cancer cells. Oncogene 18: 6322-6329. doi: 10.1038/sj.onc.1203032
|
| [38] |
Huggins C, Hodges CV (1972) Studies on prostatic cancer. I. The effect of castration, of estrogen and androgen injection on serum phosphatases in metastatic carcinoma of the prostate. CA Cancer J Clin 22: 232-240. doi: 10.3322/canjclin.22.4.232
|
| [39] |
Harris WP, Mostaghel EA, Nelson PS, et al. (2009) Androgen deprivation therapy: progress in understanding mechanisms of resistance and optimizing androgen depletion. Nat Clin Pract Urol 6: 76-85. doi: 10.1038/ncpuro1296
|
| [40] |
Klugo RC, Farah RN, Cerny JC (1981) Bilateral orchiectomy for carcinoma of prostate. Response of serum testosterone and clinical response to subsequent estrogen therapy. Urology 17: 49-50. doi: 10.1016/0090-4295(81)90011-X
|
| [41] |
Labrie F, Dupont A, Belanger A, et al. (1986) Treatment of prostate cancer with gonadotropin-releasing hormone agonists. Endocr Rev 7: 67-74. doi: 10.1210/edrv-7-1-67
|
| [42] | Labrie F, Belanger A, Dupont A, et al. (1993) Science behind total androgen blockade: from gene to combination therapy. Clin Invest Med 16: 475-492. |
| [43] |
Karantanos T, Corn PG, Thompson TC (2013) Prostate cancer progression after androgen deprivation therapy: mechanisms of castrate resistance and novel therapeutic approaches. Oncogene 32: 5501-5511. doi: 10.1038/onc.2013.206
|
| [44] |
de Bono JS, Logothetis CJ, Molina A, et al. (2011) Abiraterone and increased survival in metastatic prostate cancer. N Engl J Med 364: 1995-2005. doi: 10.1056/NEJMoa1014618
|
| [45] |
Tran C, Ouk S, Clegg NJ, et al. (2009) Development of a second-generation antiandrogen for treatment of advanced prostate cancer. Science 324: 787-790. doi: 10.1126/science.1168175
|
| [46] |
Kluetz PG, Pierce W, Maher VE, et al. (2014) Radium Ra 223 dichloride injection: U.S. Food and Drug Administration drug approval summary. Clin Cancer Res 20: 9-14. doi: 10.1158/1078-0432.CCR-13-2665
|
| [47] |
Pal S, Gupta R, Davuluri RV (2012) Alternative transcription and alternative splicing in cancer. Pharmacol Ther 136: 283-294. doi: 10.1016/j.pharmthera.2012.08.005
|
| [48] |
Heuze-Vourc'h N, Leblond V, Courty Y (2003) Complex alternative splicing of the hKLK3 gene coding for the tumor marker PSA (prostate-specific-antigen). Eur J Biochem 270: 706-714. doi: 10.1046/j.1432-1033.2003.03425.x
|
| [49] |
Narla G, DiFeo A, Fernandez Y, et al. (2008) KLF6-SV1 overexpression accelerates human and mouse prostate cancer progression and metastasis. J Clin Invest 118: 2711-2721. doi: 10.1172/JCI34780
|
| [50] |
Thorsen K, Sorensen KD, Brems-Eskildsen AS, et al. (2008) Alternative splicing in colon, bladder, and prostate cancer identified by exon array analysis. Mol Cell Proteomics 7: 1214-1224. doi: 10.1074/mcp.M700590-MCP200
|
| [51] | Erho N, Buerki C, Triche TJ, et al. (2012) Transcriptome-wide detection of differentially expressed coding and non-coding transcripts and their clinical significance in prostate cancer. J Oncol 2012: 541353. |
| [52] |
Rajan P, Dalgliesh C, Carling PJ, et al. (2011) Identification of novel androgen-regulated pathways and mRNA isoforms through genome-wide exon-specific profiling of the LNCaP transcriptome. PLoS One 6: e29088. doi: 10.1371/journal.pone.0029088
|
| [53] | Munkley J, Rajan P, Lafferty NP, et al. (2014) A novel androgen-regulated isoform of the TSC2 tumour suppressor gene increases cell proliferation. Oncotarget 5: 131-139. |
| [54] |
Busa R, Paronetto MP, Farini D, et al. (2007) The RNA-binding protein Sam68 contributes to proliferation and survival of human prostate cancer cells. Oncogene 26: 4372-4382. doi: 10.1038/sj.onc.1210224
|
| [55] |
Rajan P, Gaughan L, Dalgliesh C, et al. (2008) The RNA-binding and adaptor protein Sam68 modulates signal-dependent splicing and transcriptional activity of the androgen receptor. J Pathol 215: 67-77. doi: 10.1002/path.2324
|
| [56] | Mavrou A, Brakspear K, Hamdollah-Zadeh M, et al. (2014) Serine-arginine protein kinase 1 (SRPK1) inhibition as a potential novel targeted therapeutic strategy in prostate cancer. Oncogene 34: 4311-4319. |
| [57] |
Clark EL, Coulson A, Dalgliesh C, et al. (2008) The RNA helicase p68 is a novel androgen receptor coactivator involved in splicing and is overexpressed in prostate cancer. Cancer Res 68: 7938-7946. doi: 10.1158/0008-5472.CAN-08-0932
|
| [58] | Wu X, Daniels G, Lee P, et al. (2014) Lipid metabolism in prostate cancer. Am J Clin Exp Urol 2: 111-120. |
| [59] |
Suburu J, Chen YQ (2012) Lipids and prostate cancer. Prostaglandins Other Lipid Mediat 98: 1-10. doi: 10.1016/j.prostaglandins.2012.03.003
|
| [60] |
Swinnen JV, Ulrix W, Heyns W, et al. (1997) Coordinate regulation of lipogenic gene expression by androgens: evidence for a cascade mechanism involving sterol regulatory element binding proteins. Proc Natl Acad Sci U S A 94: 12975-12980. doi: 10.1073/pnas.94.24.12975
|
| [61] | Swinnen JV, Esquenet M, Goossens K, et al. (1997) Androgens stimulate fatty acid synthase in the human prostate cancer cell line LNCaP. Cancer Res 57: 1086-1090. |
| [62] | Swinnen JV, Vanderhoydonc F, Elgamal AA, et al. (2000) Selective activation of the fatty acid synthesis pathway in human prostate cancer. Int J Cancer 88: 176-179. |
| [63] |
Liu Y (2006) Fatty acid oxidation is a dominant bioenergetic pathway in prostate cancer. Prostate Cancer Prostatic Dis 9: 230-234. doi: 10.1038/sj.pcan.4500879
|
| [64] |
Segawa T, Nau ME, Xu LL, et al. (2002) Androgen-induced expression of endoplasmic reticulum (ER) stress response genes in prostate cancer cells. Oncogene 21: 8749-8758. doi: 10.1038/sj.onc.1205992
|
| [65] |
Sheng X, Arnoldussen YJ, Storm M, et al. (2015) Divergent androgen regulation of unfolded protein response pathways drives prostate cancer. EMBO Mol Med 7: 788-801. doi: 10.15252/emmm.201404509
|
| [66] | Munkley J, Mills IG, Elliott DJ (2016) The role of glycans in the development and progression of prostate cancer. Nat Rev Urol 13: 324-333. |
| [67] | Hakomori S (1996) Tumor malignancy defined by aberrant glycosylation and sphingo(glyco)lipid metabolism. Cancer Res 56: 5309-5318. |
| [68] |
Reis CA, Osorio H, Silva L, et al. (2010) Alterations in glycosylation as biomarkers for cancer detection. J Clin Pathol 63: 322-329. doi: 10.1136/jcp.2009.071035
|
| [69] |
Pinho SS, Reis CA (2015) Glycosylation in cancer: mechanisms and clinical implications. Nat Rev Cancer 15: 540-555. doi: 10.1038/nrc3982
|
| [70] |
Barfeld SJ, East P, Zuber V, et al. (2014) Meta-analysis of prostate cancer gene expression data identifies a novel discriminatory signature enriched for glycosylating enzymes. BMC Med Genom 7: 513. doi: 10.1186/s12920-014-0074-9
|
| [71] | Munkley J, Elliott DJ (2016) Hallmarks of glycosylation in cancer. Oncotarget in press. |
| [72] | Munkley J, Vodak D, Livermore KE, et al. (2016) Glycosylation is an androgen-regulated process essential for prostate cancer cell viability. EBioMedicine in press. |
| [73] |
Itkonen HM, Engedal N, Babaie E, et al. (2015) UAP1 is overexpressed in prostate cancer and is protective against inhibitors of N-linked glycosylation. Oncogene 34: 3744-3750. doi: 10.1038/onc.2014.307
|
| [74] | Munkley J, Oltean S, Vodak D, et al. (2015) The androgen receptor controls expression of the cancer-associated sTn antigen and cell adhesion through induction of ST6GalNAc1 in prostate cancer. Oncotarget 6: 34358-34374. |
| [75] |
Sato T, Yoneyama T, Tobisawa Y, et al. (2016) Core 2 (beta-1, 6-N-acetylglucosaminyltransferase-1 expression in prostate biopsy specimen is an indicator of prostate cancer aggressiveness. Biochem Biophys Res Commun 470: 150-156. doi: 10.1016/j.bbrc.2016.01.011
|
| [76] |
Chen Z, Gulzar ZG, St Hill CA, et al. (2014) Increased expression of GCNT1 is associated with altered O-glycosylation of PSA, PAP, and MUC1 in human prostate cancers. Prostate 74: 1059-1067. doi: 10.1002/pros.22826
|
| [77] |
Taylor BS, Schultz N, Hieronymus H, et al. (2010) Integrative genomic profiling of human prostate cancer. Cancer Cell 18: 11-22. doi: 10.1016/j.ccr.2010.05.026
|
| [78] | Pourmand G, Ziaee AA, Abedi AR, et al. (2007) Role of PTEN gene in progression of prostate cancer. Urol J 4: 95-100. |
| [79] |
Robinson D, Van Allen EM, Wu YM, et al. (2015) Integrative clinical genomics of advanced prostate cancer. Cell 161: 1215-1228. doi: 10.1016/j.cell.2015.05.001
|
| [80] |
Carver BS, Chapinski C, Wongvipat J, et al. (2011) Reciprocal feedback regulation of PI3K and androgen receptor signaling in PTEN-deficient prostate cancer. Cancer Cell 19: 575-586. doi: 10.1016/j.ccr.2011.04.008
|
| [81] |
Munkley J, Lafferty NP, Kalna G, et al. (2015) Androgen-regulation of the protein tyrosine phosphatase PTPRR activates ERK1/2 signalling in prostate cancer cells. BMC Cancer 15: 9. doi: 10.1186/s12885-015-1012-8
|
| [82] |
Schutzman JL, Martin GR (2012) Sprouty genes function in suppression of prostate tumorigenesis. Proc Natl Acad Sci U S A 109: 20023-20028. doi: 10.1073/pnas.1217204109
|
| [83] | Yokoyama NN, Shao S, Hoang BH, et al. (2014) Wnt signaling in castration-resistant prostate cancer: implications for therapy. Am J Clin Exp Urol 2: 27-44. |
| [84] | Truica CI, Byers S, Gelmann EP (2000) Beta-catenin affects androgen receptor transcriptional activity and ligand specificity. Cancer Res 60: 4709-4713. |
| [85] |
Mulholland DJ, Cheng H, Reid K, et al. (2002) The androgen receptor can promote beta-catenin nuclear translocation independently of adenomatous polyposis coli. J Biol Chem 277: 17933-17943. doi: 10.1074/jbc.M200135200
|
| [86] |
Rajan P, Sudbery IM, Villasevil ME, et al. (2014) Next-generation sequencing of advanced prostate cancer treated with androgen-deprivation therapy. Eur Urol 66: 32-39. doi: 10.1016/j.eururo.2013.08.011
|
| [87] |
Chen G, Shukeir N, Potti A, et al. (2004) Up-regulation of Wnt-1 and beta-catenin production in patients with advanced metastatic prostate carcinoma: potential pathogenetic and prognostic implications. Cancer 101: 1345-1356. doi: 10.1002/cncr.20518
|
| [88] | Voeller HJ, Truica CI, Gelmann EP (1998) Beta-catenin mutations in human prostate cancer. Cancer Res 58: 2520-2523. |
| [89] |
Mostaghel EA, Solomon KR, Pelton K, et al. (2012) Impact of circulating cholesterol levels on growth and intratumoral androgen concentration of prostate tumors. PLoS One 7: e30062. doi: 10.1371/journal.pone.0030062
|
| [90] | Hamid AR, Pfeiffer MJ, Verhaegh GW, et al. (2012) Aldo-keto reductase family 1 member C3 (AKR1C3) is a biomarker and therapeutic target for castration-resistant prostate cancer. Mol Med 18: 1449-1455. |
| [91] |
Stanbrough M, Bubley GJ, Ross K, et al. (2006) Increased expression of genes converting adrenal androgens to testosterone in androgen-independent prostate cancer. Cancer Res 66: 2815-2825. doi: 10.1158/0008-5472.CAN-05-4000
|
| [92] |
Mohler JL, Gregory CW, Ford OH, 3rd, et al. (2004) The androgen axis in recurrent prostate cancer. Clin Cancer Res 10: 440-448. doi: 10.1158/1078-0432.CCR-1146-03
|
| [93] |
Knuuttila M, Yatkin E, Kallio J, et al. (2014) Castration induces up-regulation of intratumoral androgen biosynthesis and androgen receptor expression in an orthotopic VCaP human prostate cancer xenograft model. Am J Pathol 184: 2163-2173. doi: 10.1016/j.ajpath.2014.04.010
|
| [94] |
Liu W, Xie CC, Zhu Y, et al. (2008) Homozygous deletions and recurrent amplifications implicate new genes involved in prostate cancer. Neoplasia 10: 897-907. doi: 10.1593/neo.08428
|
| [95] |
Edwards J, Krishna NS, Grigor KM, et al. (2003) Androgen receptor gene amplification and protein expression in hormone refractory prostate cancer. Br J Cancer 89: 552-556. doi: 10.1038/sj.bjc.6601127
|
| [96] | Edwards J, Krishna NS, Witton CJ, et al. (2003) Gene amplifications associated with the development of hormone-resistant prostate cancer. Clin Cancer Res 9: 5271-5281. |
| [97] |
Holzbeierlein J, Lal P, LaTulippe E, et al. (2004) Gene expression analysis of human prostate carcinoma during hormonal therapy identifies androgen-responsive genes and mechanisms of therapy resistance. Am J Pathol 164: 217-227. doi: 10.1016/S0002-9440(10)63112-4
|
| [98] |
Barbieri CE, Bangma CH, Bjartell A, et al. (2013) The mutational landscape of prostate cancer. Eur Urol 64: 567-576. doi: 10.1016/j.eururo.2013.05.029
|
| [99] |
Grasso CS, Wu YM, Robinson DR, et al. (2012) The mutational landscape of lethal castration-resistant prostate cancer. Nature 487: 239-243. doi: 10.1038/nature11125
|
| [100] |
Shaffer DR, Leversha MA, Danila DC, et al. (2007) Circulating tumor cell analysis in patients with progressive castration-resistant prostate cancer. Clin Cancer Res 13: 2023-2029. doi: 10.1158/1078-0432.CCR-06-2701
|
| [101] |
Salvi S, Casadio V, Conteduca V, et al. (2015) Circulating cell-free AR and CYP17A1 copy number variations may associate with outcome of metastatic castration-resistant prostate cancer patients treated with abiraterone. Br J Cancer 112: 1717-1724. doi: 10.1038/bjc.2015.128
|
| [102] |
Azad AA, Volik SV, Wyatt AW, et al. (2015) Androgen Receptor Gene Aberrations in Circulating Cell-Free DNA: Biomarkers of Therapeutic Resistance in Castration-Resistant Prostate Cancer. Clin Cancer Res 21: 2315-2324. doi: 10.1158/1078-0432.CCR-14-2666
|
| [103] |
Attard G, Swennenhuis JF, Olmos D, et al. (2009) Characterization of ERG, AR and PTEN gene status in circulating tumor cells from patients with castration-resistant prostate cancer. Cancer Res 69: 2912-2918. doi: 10.1158/0008-5472.CAN-08-3667
|
| [104] | Wallen MJ, Linja M, Kaartinen K, et al. (1999) Androgen receptor gene mutations in hormone-refractory prostate cancer. J Pathol 189: 559-563. |
| [105] |
Gottlieb B, Beitel LK, Nadarajah A, et al. (2012) The androgen receptor gene mutations database: 2012 update. Hum Mutat 33: 887-894. doi: 10.1002/humu.22046
|
| [106] | Gaddipati JP, McLeod DG, Heidenberg HB, et al. (1994) Frequent detection of codon 877 mutation in the androgen receptor gene in advanced prostate cancers. Cancer Res 54: 2861-2864. |
| [107] |
Steketee K, Timmerman L, Ziel-van der Made AC, et al. (2002) Broadened ligand responsiveness of androgen receptor mutants obtained by random amino acid substitution of H874 and mutation hot spot T877 in prostate cancer. Int J Cancer 100: 309-317. doi: 10.1002/ijc.10495
|
| [108] |
Steinkamp MP, O'Mahony OA, Brogley M, et al. (2009) Treatment-dependent androgen receptor mutations in prostate cancer exploit multiple mechanisms to evade therapy. Cancer Res 69: 4434-4442. doi: 10.1158/0008-5472.CAN-08-3605
|
| [109] | Hu R, Dunn TA, Wei S, et al. (2009) Ligand-independent androgen receptor variants derived from splicing of cryptic exons signify hormone-refractory prostate cancer. Cancer Res 69: 16-22. |
| [110] |
Sun S, Sprenger CC, Vessella RL, et al. (2010) Castration resistance in human prostate cancer is conferred by a frequently occurring androgen receptor splice variant. J Clin Invest 120: 2715-2730. doi: 10.1172/JCI41824
|
| [111] |
Hu R, Lu C, Mostaghel EA, et al. (2012) Distinct transcriptional programs mediated by the ligand-dependent full-length androgen receptor and its splice variants in castration-resistant prostate cancer. Cancer Res 72: 3457-3462. doi: 10.1158/0008-5472.CAN-11-3892
|
| [112] |
Li Y, Chan SC, Brand LJ, et al. (2013) Androgen receptor splice variants mediate enzalutamide resistance in castration-resistant prostate cancer cell lines. Cancer Res 73: 483-489. doi: 10.1158/0008-5472.CAN-12-3630
|
| [113] |
Hornberg E, Ylitalo EB, Crnalic S, et al. (2011) Expression of androgen receptor splice variants in prostate cancer bone metastases is associated with castration-resistance and short survival. PLoS One 6: e19059. doi: 10.1371/journal.pone.0019059
|
| [114] |
Antonarakis ES, Lu C, Wang H, et al. (2014) AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N Engl J Med 371: 1028-1038. doi: 10.1056/NEJMoa1315815
|
| [115] | Steinestel J, Luedeke M, Arndt A, et al. (2015) Detecting predictive androgen receptor modifications in circulating prostate cancer cells. Oncotarget 5. |
| [116] |
Culig Z (2016) Androgen Receptor Coactivators in Regulation of Growth and Differentiation in Prostate Cancer. J Cell Physiol 231: 270-274. doi: 10.1002/jcp.25099
|
| [117] |
Heemers HV, Sebo TJ, Debes JD, et al. (2007) Androgen deprivation increases p300 expression in prostate cancer cells. Cancer Res 67: 3422-3430. doi: 10.1158/0008-5472.CAN-06-2836
|
| [118] |
Comuzzi B, Nemes C, Schmidt S, et al. (2004) The androgen receptor co-activator CBP is up-regulated following androgen withdrawal and is highly expressed in advanced prostate cancer. J Pathol 204: 159-166. doi: 10.1002/path.1609
|
| [119] |
Halkidou K, Gnanapragasam VJ, Mehta PB, et al. (2003) Expression of Tip60, an androgen receptor coactivator, and its role in prostate cancer development. Oncogene 22: 2466-2477. doi: 10.1038/sj.onc.1206342
|
| [120] | Debes JD, Schmidt LJ, Huang H, et al. (2002) p300 mediates androgen-independent transactivation of the androgen receptor by interleukin 6. Cancer Res 62: 5632-5636. |
| [121] | Shiota M, Yokomizo A, Masubuchi D, et al. (2010) Tip60 Promotes Prostate Cancer Cell Proliferation by Translocation of Androgen Receptor into the Nucleus. Prostate 70: 540-554. |
| [122] | Kishimoto T (1989) The biology of interleukin-6. Blood 74: 1-10. |
| [123] |
Okamoto M, Lee C, Oyasu R (1997) Autocrine effect of androgen on proliferation of an androgen responsive prostatic carcinoma cell line, LNCAP: role of interleukin-6. Endocrinology 138: 5071-5074. doi: 10.1210/endo.138.11.5653
|
| [124] | Hobisch A, Rogatsch H, Hittmair A, et al. (2000) Immunohistochemical localization of interleukin-6 and its receptor in benign, premalignant and malignant prostate tissue. J Pathol 191: 239-244. |
| [125] |
Culig Z, Bartsch G, Hobisch A (2002) Interleukin-6 regulates androgen receptor activity and prostate cancer cell growth. Mol Cell Endocrinol 197: 231-238. doi: 10.1016/S0303-7207(02)00263-0
|
| [126] |
Twillie DA, Eisenberger MA, Carducci MA, et al. (1995) Interleukin-6: a candidate mediator of human prostate cancer morbidity. Urology 45: 542-549. doi: 10.1016/S0090-4295(99)80034-X
|
| [127] | Chen T, Wang LH, Farrar WL (2000) Interleukin 6 activates androgen receptor-mediated gene expression through a signal transducer and activator of transcription 3-dependent pathway in LNCaP prostate cancer cells. Cancer Res 60: 2132-2135. |
| [128] |
Heinrich PC, Behrmann I, Haan S, et al. (2003) Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem J 374: 1-20. doi: 10.1042/bj20030407
|
| [129] | Olayioye MA, Neve RM, Lane HA, et al. (2000) The ErbB signaling network: receptor heterodimerization in development and cancer. EMBO J 19: 3159-3167. |
| [130] |
Slamon DJ, Godolphin W, Jones LA, et al. (1989) Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science 244: 707-712. doi: 10.1126/science.2470152
|
| [131] | Signoretti S, Montironi R, Manola J, et al. (2000) Her-2-neu expression and progression toward androgen independence in human prostate cancer. J Natl Cancer Inst 92: 1918-1925. |
| [132] | Wen Y, Hu MC, Makino K, et al. (2000) HER-2/neu promotes androgen-independent survival and growth of prostate cancer cells through the Akt pathway. Cancer Res 60: 6841-6845. |
| [133] |
Karin M (2006) Nuclear factor-kappaB in cancer development and progression. Nature 441: 431-436. doi: 10.1038/nature04870
|
| [134] |
Suh J, Payvandi F, Edelstein LC, et al. (2002) Mechanisms of constitutive NF-kappaB activation in human prostate cancer cells. Prostate 52: 183-200. doi: 10.1002/pros.10082
|
| [135] |
Zhang L, Altuwaijri S, Deng F, et al. (2009) NF-kappaB regulates androgen receptor expression and prostate cancer growth. Am J Pathol 175: 489-499. doi: 10.2353/ajpath.2009.080727
|
| [136] |
Nadiminty N, Lou W, Sun M, et al. (2010) Aberrant activation of the androgen receptor by NF-kappaB2/p52 in prostate cancer cells. Cancer Res 70: 3309-3319. doi: 10.1158/0008-5472.CAN-09-3703
|
| [137] |
Krijnen JL, Janssen PJ, Ruizeveld de Winter JA, et al. (1993) Do neuroendocrine cells in human prostate cancer express androgen receptor? Histochemistry 100: 393-398. doi: 10.1007/BF00268938
|
| [138] |
Bonkhoff H, Stein U, Remberger K (1995) Endocrine-paracrine cell types in the prostate and prostatic adenocarcinoma are postmitotic cells. Hum Pathol 26: 167-170. doi: 10.1016/0046-8177(95)90033-0
|
| [139] | Li Q, Zhang CS, Zhang Y (2016) Molecular aspects of prostate cancer with neuroendocrine differentiation. Chin J Cancer Res 28: 122-129. |
| [140] | Alberti C (2010) Neuroendocrine differentiation in prostate carcinoma: focusing on its pathophysiologic mechanisms and pathological features. G Chir 31: 568-574. |
| [141] |
Yuan TC, Veeramani S, Lin FF, et al. (2006) Androgen deprivation induces human prostate epithelial neuroendocrine differentiation of androgen-sensitive LNCaP cells. Endocr Relat Cancer 13: 151-167. doi: 10.1677/erc.1.01043
|
| [142] |
Jongsma J, Oomen MH, Noordzij MA, et al. (1999) Kinetics of neuroendocrine differentiation in an androgen-dependent human prostate xenograft model. Am J Pathol 154: 543-551. doi: 10.1016/S0002-9440(10)65300-X
|
| [143] |
Hirano D, Okada Y, Minei S, et al. (2004) Neuroendocrine differentiation in hormone refractory prostate cancer following androgen deprivation therapy. Eur Urol 45: 586-592. doi: 10.1016/j.eururo.2003.11.032
|
| [144] |
Sagnak L, Topaloglu H, Ozok U, et al. (2011) Prognostic significance of neuroendocrine differentiation in prostate adenocarcinoma. Clin Genitourin Cancer 9: 73-80. doi: 10.1016/j.clgc.2011.07.003
|
| [145] |
Qiu Y, Robinson D, Pretlow TG, et al. (1998) Etk/Bmx, a tyrosine kinase with a pleckstrin-homology domain, is an effector of phosphatidylinositol 3'-kinase and is involved in interleukin 6-induced neuroendocrine differentiation of prostate cancer cells. Proc Natl Acad Sci U S A 95: 3644-3649. doi: 10.1073/pnas.95.7.3644
|
| [146] | Spiotto MT, Chung TD (2000) STAT3 mediates IL-6-induced neuroendocrine differentiation in prostate cancer cells. Prostate 42: 186-195. |
| [147] | Moritz T, Venz S, Junker H, et al. (2016) Isoform 1 of TPD52 (PC-1) promotes neuroendocrine transdifferentiation in prostate cancer cells. Tumour Biol in press. |
| [148] | Gururajan M, Cavassani KA, Sievert M, et al. (2015) SRC family kinase FYN promotes the neuroendocrine phenotype and visceral metastasis in advanced prostate cancer. Oncotarget 6: 44072-44083. |
| [149] |
Uysal-Onganer P, Kawano Y, Caro M, et al. (2010) Wnt-11 promotes neuroendocrine-like differentiation, survival and migration of prostate cancer cells. Mol Cancer 9: 55. doi: 10.1186/1476-4598-9-55
|
| [150] | Wu C, Huang J (2007) Phosphatidylinositol 3-kinase-AKT-mammalian target of rapamycin pathway is essential for neuroendocrine differentiation of prostate cancer. J Biol Chem 282: 3571-3583. |
| [151] |
Kowalska K, Piastowska-Ciesielska AW (2016) Oestrogens and oestrogen receptors in prostate cancer. Springerplus 5: 522. doi: 10.1186/s40064-016-2185-6
|
| [152] |
Thompson VC, Morris TG, Cochrane DR, et al. (2006) Relaxin becomes upregulated during prostate cancer progression to androgen independence and is negatively regulated by androgens. Prostate 66: 1698-1709. doi: 10.1002/pros.20423
|
| [153] |
Liu S, Vinall RL, Tepper C, et al. (2008) Inappropriate activation of androgen receptor by relaxin via beta-catenin pathway. Oncogene 27: 499-505. doi: 10.1038/sj.onc.1210671
|
| [154] |
Yu S, Xia S, Yang D, et al. (2013) Androgen receptor in human prostate cancer-associated fibroblasts promotes prostate cancer epithelial cell growth and invasion. Med Oncol 30: 674. doi: 10.1007/s12032-013-0674-9
|
| [155] |
Roberts AB, Anzano MA, Wakefield LM, et al. (1985) Type beta transforming growth factor: a bifunctional regulator of cellular growth. Proc Natl Acad Sci U S A 82: 119-123. doi: 10.1073/pnas.82.1.119
|
| [156] |
Fuzio P, Ditonno P, Rutigliano M, et al. (2012) Regulation of TGF-beta1 expression by androgen deprivation therapy of prostate cancer. Cancer Lett 318: 135-144. doi: 10.1016/j.canlet.2011.08.034
|
| [157] |
Yang F, Tuxhorn JA, Ressler SJ, et al. (2005) Stromal expression of connective tissue growth factor promotes angiogenesis and prostate cancer tumorigenesis. Cancer Res 65: 8887-8895. doi: 10.1158/0008-5472.CAN-05-1702
|
| [158] |
Yang F, Chen Y, Shen T, et al. (2014) Stromal TGF-beta signaling induces AR activation in prostate cancer. Oncotarget 5: 10854-10869. doi: 10.18632/oncotarget.2536
|
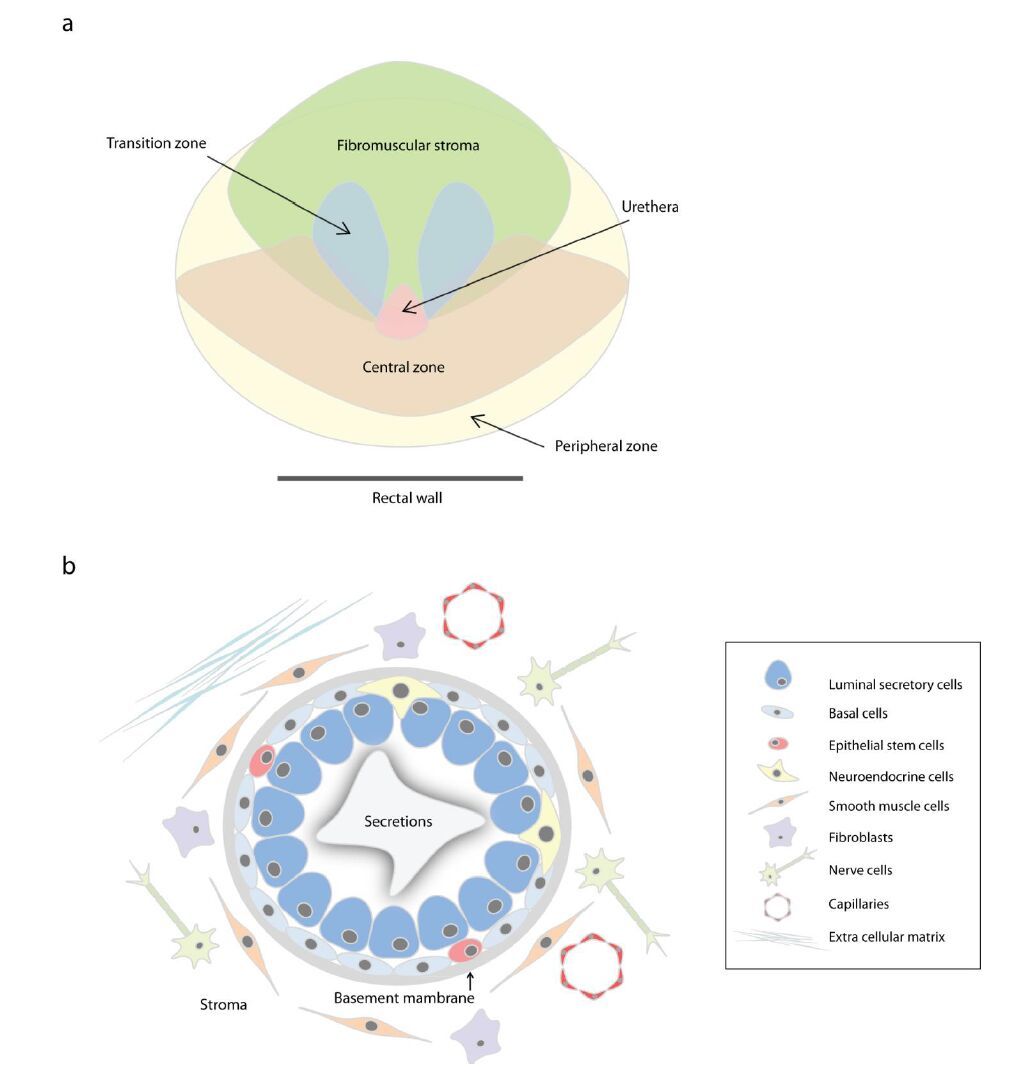

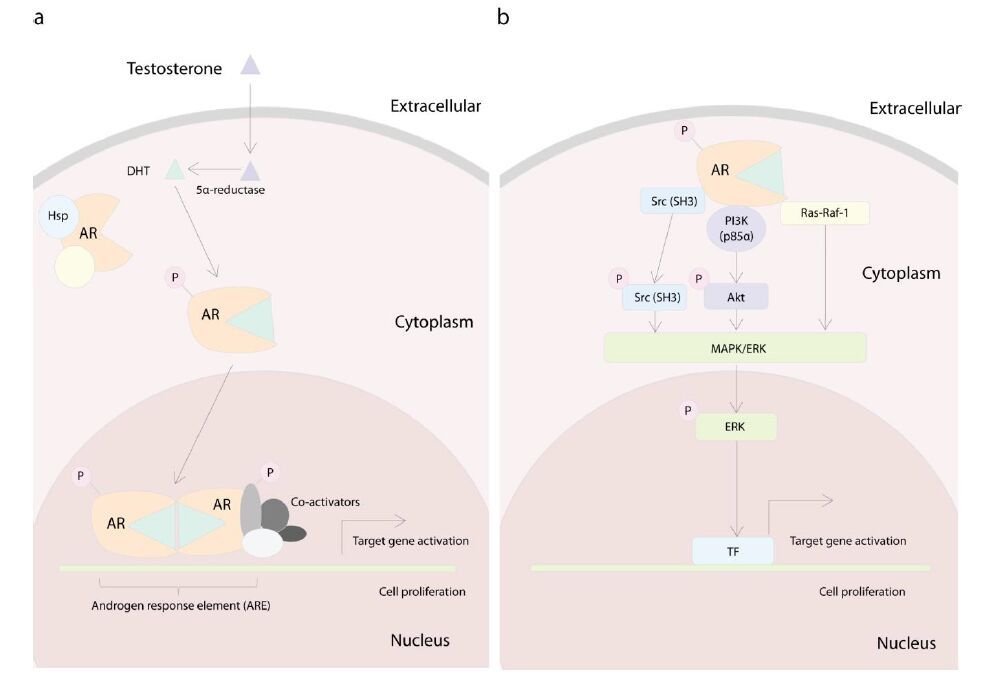
Figures(3)

Karen E. Livermore, Jennifer Munkley, David J. Elliott. Androgen receptor and prostate cancer[J]. AIMS Molecular Science, 2016, 3(2): 280-299. doi: 10.3934/molsci.2016.2.280







 DownLoad:
DownLoad: