Citation: José Rodrigo da-Silva, Ana Alexandre, Clarisse Brígido, Solange Oliveira. Can stress response genes be used to improve the symbiotic performance of rhizobia?[J]. AIMS Microbiology, 2017, 3(3): 365-382. doi: 10.3934/microbiol.2017.3.365

| [1] | Hawkesford M, Horst W, Kichey T, et al. (2012) Chapter 6-Functions of Macronutrients A2- Marschner, In: Marschner's Mineral Nutrition of Higher Plants, 3Eds., San Diego: Academic Press, 135–189. |
| [2] | Vance CP (2001) Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resources. Plant Physiol 127: 390–397. |
| [3] |
Richardson AE, Barea JM, McNeill AM, et al. (2009) Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 321: 305–339. doi: 10.1007/s11104-009-9895-2

|
| [4] |
Jensen ES, Peoples MB, Boddey RM, et al. (2012) Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agron Sustain Dev 32: 329–364. doi: 10.1007/s13593-011-0056-7
|
| [5] |
Philippot L, Hallin S (2011) Towards food, feed and energy crops mitigating climate change. Trends Plant Sci 16: 476–480. doi: 10.1016/j.tplants.2011.05.007
|
| [6] |
Oldroyd GED (2013) Speak, friend, and enter: signalling systems that promote beneficial symbiotic associations in plants. Nat Rev Microbiol 11: 252–263. doi: 10.1038/nrmicro2990
|
| [7] |
Perret X, Staehelin C, Broughton WJ (2000) Molecular basis of symbiotic promiscuity. Microbiol Mol Biol Rev 64: 180–201. doi: 10.1128/MMBR.64.1.180-201.2000
|
| [8] | Downie JA (1998) Functions of rhizobial nodulation genes, In: Spaink HP, Kondorosi A, Hooykaas PJJ, Editors, The Rhizobiaceae Molecular Biology of Model Plant-Associated Bacteria, Dordrecht: Kluwer Academic Publishers, 387–402. |
| [9] |
Oldroyd GE, Murray JD, Poole PS, et al. (2011) The rules of engagement in the legume-rhizobial symbiosis. Annu Rev Genet 45: 119–144. doi: 10.1146/annurev-genet-110410-132549
|
| [10] | Sprent JI (2009) Evolution of Nodulation, In: Legume Nodulation, Wiley-Blackwell, 51–63. |
| [11] |
Oldroyd GED, Downie JM (2008) Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu Rev Plant Biol 59: 519–546. doi: 10.1146/annurev.arplant.59.032607.092839
|
| [12] |
Chen WM, Moulin L, Bontemps C, et al. (2003) Legume symbiotic nitrogen fixation by β-proteobacteria is widespread in nature. J Bacteriol 185: 7266–7272. doi: 10.1128/JB.185.24.7266-7272.2003
|
| [13] |
Moulin L, Klonowska A, Caroline B, et al. (2014) Complete genome sequence of Burkholderia phymatum STM815(T), a broad host range and efficient nitrogen-fixing symbiont of Mimosa species. Stand Genomic Sci 9: 763–774. doi: 10.4056/sigs.4861021
|
| [14] |
Peix A, Ramírez-Bahena MH, Velázquez E, et al. (2015) Bacterial associations with legumes. Crit Rev Plant Sci 34: 17–42. doi: 10.1080/07352689.2014.897899
|
| [15] | Alexandre A, Oliveira S (2016) Heat shock response in bacteria with large genomes: lessons from rhizobia, In: de Bruijn FJ, Editor, Stress and Environmental Control of Gene Expression in Bacteria, Wiley-Blackwell Publishers, 737–746. |
| [16] |
Crossman LC, Castillo-Ramírez S, McAnnula C, et al. (2008) A common genomic framework for a diverse assembly of plasmids in the symbiotic nitrogen fixing bacteria. PLoS One 3: e2567. doi: 10.1371/journal.pone.0002567
|
| [17] |
Young JP, Crossman L, Johnston A, et al. (2006) The genome of Rhizobium leguminosarum has recognizable core and accessory components. Genome Biol 7: R34. doi: 10.1186/gb-2006-7-4-r34
|
| [18] |
MacLean AM, Finan TM, Sadowsky MJ (2007) Genomes of the symbiotic nitrogen-fixing bacteria of legumes. Plant Physiol 144: 615–622. doi: 10.1104/pp.107.101634
|
| [19] |
Laranjo M, Alexandre A, Oliveira S (2014) Legume growth-promoting rhizobia: an overview on the Mesorhizobium genus. Microbiol Res 169: 2–17. doi: 10.1016/j.micres.2013.09.012
|
| [20] |
Via VD, Zanetti ME, Blanco F (2016) How legumes recognize rhizobia. Plant Signal Behav 11: e1120396. doi: 10.1080/15592324.2015.1120396
|
| [21] |
Limpens E, van Zeijl A, Geurts R (2015) Lipochitooligosaccharides modulate plant host immunity to enable endosymbioses. Annu Rev Phytopathol 53: 311–334. doi: 10.1146/annurev-phyto-080614-120149
|
| [22] | Kaminski PA, Batut J, Boistard P (1998) A Survey of Symbiotic Nitrogen Fixation by Rhizobia, In: Spaink HP, Kondorosi A, Hooykaas PJJ, Editors, The Rhizobiaceae: Molecular Biology of Model Plant-Associated Bacteria, Dordrecht: Kluwer Academic Publishers, 432–460. |
| [23] |
Tschauner K, Hornschemeyer P, Muller VS, et al. (2014) Dynamic interaction between the CpxA sensor kinase and the periplasmic accessory protein CpxP mediates signal recognition in E. coli. PLoS One 9: e107383. doi: 10.1371/journal.pone.0107383
|
| [24] |
Narberhaus F (2010) Translational control of bacterial heat shock and virulence genes by temperature-sensing mRNAs. RNA Biol 7: 84–89. doi: 10.4161/rna.7.1.10501
|
| [25] |
Feklistov A, Sharon BD, Darst SA, et al. (2014) Bacterial sigma factors: a historical, structural, and genomic perspective. Annu Rev Microbiol 68: 357–376. doi: 10.1146/annurev-micro-092412-155737
|
| [26] | Nonaka G, Blankschien M, Herman C, et al. (2006) Regulon and promoter analysis of the E. coli heat-shock factor, sigma32, reveals a multifaceted cellular response to heat stress. Genes Dev 20: 1776–1789. |
| [27] |
Schumann W (2016) Regulation of bacterial heat shock stimulons. Cell Stress Chaperon 21: 959–968. doi: 10.1007/s12192-016-0727-z
|
| [28] | Ladeiro B (2012) Saline agriculture in the 21st century: using salt contaminated resources to cope food requirements. J Bot 2012: 1–7. |
| [29] | Bruning B, van Logtestijn R, Broekman R, et al. (2015) Growth and nitrogen fixation of legumes at increased salinity under field conditions: implications for the use of green manures in saline environments. AoB Plants 7. |
| [30] |
Martinez-Hidalgo P, Ramirez-Bahena MH, Flores-Felix JD, et al. (2016) Reclassification of strains MAFF 303099T and R7A into Mesorhizobium japonicum sp. nov. Int J Syst Evol Microbiol 66: 4936–4941. doi: 10.1099/ijsem.0.001448
|
| [31] |
Laranjo M, Alexandre A, Oliveira S (2017) Global transcriptional response to salt shock of the plant microsymbiont Mesorhizobium loti MAFF303099. Res Microbiol 168: 55–63. doi: 10.1016/j.resmic.2016.07.006
|
| [32] |
Lopez-Leal G, Tabche ML, Castillo-Ramirez S, et al. (2014) RNA-Seq analysis of the multipartite genome of Rhizobium etli CE3 shows different replicon contributions under heat and saline shock. BMC Genomics 15: 770. doi: 10.1186/1471-2164-15-770
|
| [33] |
Domínguez-Ferreras A, Perez-Arnedo R, Becker A, et al. (2006) Transcriptome profiling reveals the importance of plasmid pSymB for osmoadaptation of Sinorhizobium meliloti. J Bacteriol 188: 7617–7625. doi: 10.1128/JB.00719-06
|
| [34] |
Perez-Montano F, Del Cerro P, Jimenez-Guerrero I, et al. (2016) RNA-seq analysis of the Rhizobium tropici CIAT 899 transcriptome shows similarities in the activation patterns of symbiotic genes in the presence of apigenin and salt. BMC Genomics 17: 198. doi: 10.1186/s12864-016-2543-3
|
| [35] |
Liu X, Luo Y, Mohamed OA, et al. (2014) Global transcriptome analysis of Mesorhizobium alhagi CCNWXJ12-2 under salt stress. BMC Microbiol 14: 1. doi: 10.1186/1471-2180-14-1
|
| [36] |
Alexandre A, Laranjo M, Oliveira S (2014) Global transcriptional response to heat shock of the legume symbiont Mesorhizobium loti MAFF303099 comprises extensive gene downregulation. DNA Res 21: 195–206. doi: 10.1093/dnares/dst050
|
| [37] |
Barnett MJ, Bittner AN, Toman CJ, et al. (2012) Dual RpoH sigma factors and transcriptional plasticity in a symbiotic bacterium. J Bacteriol 194: 4983–4994. doi: 10.1128/JB.00449-12
|
| [38] |
Sauviac L, Philippe H, Phok K, et al. (2007) An extracytoplasmic function sigma factor acts as a general stress response regulator in Sinorhizobium meliloti. J Bacteriol 189: 4204–4216. doi: 10.1128/JB.00175-07
|
| [39] |
Laranjo M, Alexandre A, Oliveira S (2014) Genes commonly involved in acid tolerance are not overexpressed in the plant microsymbiont Mesorhizobium loti MAFF303099 upon acidic shock. Appl Microbiol Biotechnol 98: 7137–7147. doi: 10.1007/s00253-014-5875-4
|
| [40] |
Hellweg C, Puhler A, Weidner S (2009) The time course of the transcriptomic response of Sinorhizobium meliloti 1021 following a shift to acidic pH. BMC Microbiol 9: 37. doi: 10.1186/1471-2180-9-37
|
| [41] |
de Lucena DK, Puhler A, Weidner S (2010) The role of sigma factor RpoH1 in the pH stress response of Sinorhizobium meliloti. BMC Microbiol 10: 265. doi: 10.1186/1471-2180-10-265
|
| [42] |
Draghi WO, Del Papa MF, Hellweg C, et al. (2016) A consolidated analysis of the physiologic and molecular responses induced under acid stress in the legume-symbiont model-soil bacterium Sinorhizobium meliloti. Sci Rep 6: 29278. doi: 10.1038/srep29278
|
| [43] |
Fayet O, Ziegelhoffer T, Georgopoulos C (1989) The groES and groEL heat shock gene products of Escherichia coli are essential for bacterial growth at all temperatures. J Bacteriol 171: 1379–1385. doi: 10.1128/jb.171.3.1379-1385.1989
|
| [44] |
Ferrer M, Chernikova TN, Yakimov MM, et al. (2003) Chaperonins govern growth of Escherichia coli at low temperatures. Nat Biotechnol 21: 1266–1267. doi: 10.1038/nbt1103-1266
|
| [45] |
Alexandre A, Oliveira S (2013) Response to temperature stress in rhizobia. Crit Rev Microbiol 39: 219–228. doi: 10.3109/1040841X.2012.702097
|
| [46] | Fischer HM, Babst M, Kaspar T, et al. (1993) One member of a groESL-like chaperonin multigene family in Bradyrhizobium japonicum is co-regulated with symbiotic nitrogen fixation genes. EMBO J 12: 2901–2912. |
| [47] |
Bittner AN, Foltz A, Oke V (2007) Only one of five groEL genes is required for viability and successful symbiosis in Sinorhizobium meliloti. J Bacteriol 189: 1884–1889. doi: 10.1128/JB.01542-06
|
| [48] |
Rodriguez-Quinones F, Maguire M, Wallington EJ, et al. (2005) Two of the three groEL homologues in Rhizobium leguminosarum are dispensable for normal growth. Arch Microbiol 183: 253–265. doi: 10.1007/s00203-005-0768-7
|
| [49] |
Minder AC, Narberhaus F, Babst M, et al. (1997) The dnaKJ operon belongs to the sigma(32)-dependent class of heat shock genes in Bradyrhizobium japonicum. Mol Gen Genet 254: 195–206. doi: 10.1007/s004380050408
|
| [50] |
Nogales J, Campos R, BenAbdelkhalek H, et al. (2002) Rhizobium tropici genes involved in free-living salt tolerance are required for the establishment of efficient nitrogen-fixing symbiosis with Phaseolus vulgaris. Mol Plant Microbe In 15: 225–232. doi: 10.1094/MPMI.2002.15.3.225
|
| [51] |
Alexandre A, Oliveira S (2011) Most heat-tolerant rhizobia show high induction of major chaperone genes upon stress. FEMS Microbiol Ecol 75: 28–36. doi: 10.1111/j.1574-6941.2010.00993.x
|
| [52] |
Brígido C, Oliveira S (2013) Most acid-tolerant chickpea mesorhizobia show induction of major chaperone genes upon acid shock. Microb Ecol 65: 145–153. doi: 10.1007/s00248-012-0098-7
|
| [53] |
Brígido C, Alexandre A, Oliveira S (2012) Transcriptional analysis of major chaperone genes in salt-tolerant and salt-sensitive mesorhizobia. Microbiol Res 167: 623–629. doi: 10.1016/j.micres.2012.01.006
|
| [54] |
Brígido C, Robledo M, Menendez E, et al. (2012) A ClpB chaperone knockout mutant of Mesorhizobium ciceri shows a delay in the root nodulation of chickpea plants. Mol Plant Microbe In 25: 1594–1604. doi: 10.1094/MPMI-05-12-0140-R
|
| [55] | Schroth G, Lehmann J, Barrios E (2002) Soil nutrient availability and acidity, In: Schroth G, Sinclair, F. L, Editor, Trees, crops and soil fertility: concepts and research methods, CABI, 93–130. |
| [56] |
Tiwari RP, Reeve WG, Dilworth MJ, et al. (1996) An essential role for actA in acid tolerance of Rhizobium meliloti. Microbiology 142: 601–610. doi: 10.1099/13500872-142-3-601
|
| [57] |
Tiwari RP, Reeve WG, Dilworth MJ, et al. (1996) Acid tolerance in Rhizobium meliloti strain WSM419 involves a two-component sensor-regulator system. Microbiology 142: 1693–1704. doi: 10.1099/13500872-142-7-1693
|
| [58] |
Reeve WG, Brau L, Castelli J, et al. (2006) The Sinorhizobium medicae WSM419 IpiA gene is transcriptionally activated by FsrR and required to enhance survival in lethal acid conditions. Microbiology 152: 3049–3059. doi: 10.1099/mic.0.28764-0
|
| [59] |
Reeve WG, Tiwari RP, Wong CM, et al. (1998) The transcriptional regulator gene phrR in Sinorhizobium meliloti WSM419 is regulated by low pH and other stresses. Microbiology 144: 3335–3342. doi: 10.1099/00221287-144-12-3335
|
| [60] |
Reina-Bueno M, Argandona M, Nieto JJ, et al. (2012) Role of trehalose in heat and desiccation tolerance in the soil bacterium Rhizobium etli. BMC Microbiol 12: 207. doi: 10.1186/1471-2180-12-207
|
| [61] |
Moussaid S, Domínguez-Ferreras A, Muñoz S, et al. (2015) Increased trehalose biosynthesis improves Mesorhizobium ciceri growth and symbiosis establishment in saline conditions. Symbiosis 67: 103–111. doi: 10.1007/s13199-015-0338-y
|
| [62] |
Gage DJ (2004) Infection and invasion of roots by symbiotic, nitrogen-fixing rhizobia during nodulation of temperate legumes. Microbiol Mol Biol Rev 68: 280–300. doi: 10.1128/MMBR.68.2.280-300.2004
|
| [63] |
Fournier J, Timmers ACJ, Sieberer BJ, et al. (2008) Mechanism of infection thread elongation in root hairs of Medicago truncatula and dynamic interplay with associated rhizobial colonization. Plant Physiol 148: 1985–1995. doi: 10.1104/pp.108.125674
|
| [64] | Zhang F, Smith DL (1996) Genistein accumulation in soybean (Glycine max [L.] Merr.) root systems under suboptimal root zone temperatures. J Exp Bot 47: 785–792. |
| [65] |
Hungria M, Stacey G (1997) Molecular signals exchanged between host plants and rhizobia: Basic aspects and potential application in agriculture. Soil Biol Biochem 29: 819–830. doi: 10.1016/S0038-0717(96)00239-8
|
| [66] |
Hungria M, Vargas MAT (2000) Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Res 65: 151–164. doi: 10.1016/S0378-4290(99)00084-2
|
| [67] | Zahran HH (1999) Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol Mol Biol Rev 63: 968–989. |
| [68] |
Soto MJ, Dominguez-Ferreras A, Perez-Mendoza D, et al. (2009) Mutualism versus pathogenesis: the give-and-take in plant-bacteria interactions. Cellular Microbiol 11: 381–388. doi: 10.1111/j.1462-5822.2009.01282.x
|
| [69] | Marschner H (1995) Mineral nutrition of higher plants, London: Academic Press, 889. |
| [70] |
Perez-Galdona R, Kahn ML (1994) Effects of organic-acids and low pH on Rhizobium meliloti 104A14. Microbiology 140: 1231–1235. doi: 10.1099/13500872-140-5-1231
|
| [71] |
Oke V, Long SR (1999) Bacteroid formation in the Rhizobium-legume symbiosis. Curr Opin Microbiol 2: 641–646. doi: 10.1016/S1369-5274(99)00035-1
|
| [72] |
Djordjevic MA, Chen HC, Natera S, et al. (2003) A global analysis of protein expression profiles in Sinorhizobium meliloti: discovery of new genes for nodule occupancy and stress adaptation. Mol Plant Microbe In 16: 508–524. doi: 10.1094/MPMI.2003.16.6.508
|
| [73] |
Djordjevic MA (2004) Sinorhizobium meliloti metabolism in the root nodule: A proteomic perspective. Proteomics 4: 1859–1872. doi: 10.1002/pmic.200300802
|
| [74] |
Sarma AD, Emerich DW (2005) Global protein expression pattern of Bradyrhizobium japonicum bacteroids: A prelude to functional proteomics. Proteomics 5: 4170–4184. doi: 10.1002/pmic.200401296
|
| [75] |
Sarma AD, Emerich DW (2006) A comparative proteomic evaluation of culture grown vs nodule isolated Bradyrhizobium japonicum. Proteomics 6: 3008–3028. doi: 10.1002/pmic.200500783
|
| [76] |
Nomura M, Arunothayanan H, Dao TV, et al. (2010) Differential protein profiles of Bradyrhizobium japonicum USDA110 bacteroid during soybean nodule development. Soil Sci Plant Nutr 56: 579–590. doi: 10.1111/j.1747-0765.2010.00500.x
|
| [77] |
Uchiumi T, Ohwada T, Itakura M, et al. (2004) Expression islands clustered on the symbiosis island of the Mesorhizobium loti genome. J Bacteriol 186: 2439–2448. doi: 10.1128/JB.186.8.2439-2448.2004
|
| [78] |
Pessi G, Ahrens CH, Rehrauer H, et al. (2007) Genome-wide transcript analysis of Bradyrhizobium japonicum bacteroids in soybean root nodules. Mol Plant Microbe In 20: 1353–1363. doi: 10.1094/MPMI-20-11-1353
|
| [79] |
Karunakaran R, Ramachandran VK, Seaman JC, et al. (2009) Transcriptomic analysis of Rhizobium leguminosarum biovar viciae in symbiosis with host plants Pisum sativum and Vicia cracca. J Bacteriol 191: 4002–4014. doi: 10.1128/JB.00165-09
|
| [80] |
Fischer HM, Schneider K, Babst M, et al. (1999) GroEL chaperonins are required for the formation of a functional nitrogenase in Bradyrhizobium japonicum. Arch Microbiol 171: 279–289. doi: 10.1007/s002030050711
|
| [81] |
Ogawa J, Long SR (1995) The Rhizobium meliloti groELc locus is required for regulation of early nod genes by the transcription activator nodD. Genes Dev 9: 714–729. doi: 10.1101/gad.9.6.714
|
| [82] | Labidi M, Laberge S, Vezina LP, et al. (2000) The dnaJ (hsp40) locus in Rhizobium leguminosarum bv. phaseoli is required for the establishment of an effective symbiosis with Phaseolus vulgaris. Mol Plant Microbe In 13: 1271–1274. |
| [83] |
Summers ML, Elkins JG, Elliott BA, et al. (1998) Expression and regulation of phosphate stress inducible genes in Sinorhizobium meliloti. Mol Plant Microbe In 11: 1094–1101. doi: 10.1094/MPMI.1998.11.11.1094
|
| [84] |
Mitsui H, Sato T, Sato Y, et al. (2004) Sinorhizobium meliloti RpoH(1) is required for effective nitrogen-fixing symbiosis with alfalfa. Mol Genet Genomics 271: 416–425. doi: 10.1007/s00438-004-0992-x
|
| [85] |
Oke V, Rushing BG, Fisher EJ, et al. (2001) Identification of the heat-shock sigma factor RpoH and a second RpoH-like protein in Sinorhizobium meliloti. Microbiology 147: 2399–2408. doi: 10.1099/00221287-147-9-2399
|
| [86] |
Ono Y, Mitsui H, Sato T, et al. (2001) Two RpoH homologs responsible for the expression of heat shock protein genes in Sinorhizobium meliloti. Mol Gen Genet 264: 902–912. doi: 10.1007/s004380000380
|
| [87] |
Bittner AN, Oke V (2006) Multiple groESL operons are not key targets of RpoH1 and RpoH2 in Sinorhizobium meliloti. J Bacteriol 188: 3507–3515. doi: 10.1128/JB.188.10.3507-3515.2006
|
| [88] |
Martínez-Salazar JM, Sandoval-Calderon M, Guo XW, et al. (2009) The Rhizobium etli RpoH1 and RpoH2 sigma factors are involved in different stress responses. Microbiology 155: 386–397. doi: 10.1099/mic.0.021428-0
|
| [89] |
de Lucena DK, Pühler A, Weidner S (2010) The role of sigma factor RpoH1 in the pH stress response of Sinorhizobium meliloti. BMC Microbiol 10: 265. doi: 10.1186/1471-2180-10-265
|
| [90] |
Puskas LG, Nagy ZB, Kelemen JZ, et al. (2004) Wide-range transcriptional modulating effect of ntrR under microaerobiosis in Sinorhizobium meliloti. Mol Genet Genomics 272: 275–289. doi: 10.1007/s00438-004-1051-3
|
| [91] |
Martínez-Salazar JM, Sandoval-Calderon M, Guo X, et al. (2009) The Rhizobium etli RpoH1 and RpoH2 sigma factors are involved in different stress responses. Microbiology 155: 386–397. doi: 10.1099/mic.0.021428-0
|
| [92] | Martínez-Salazar JM, Salazar E, Encarnación S, et al. (2009b) Role of the extracytoplasmic function sigma factor RpoE4 in oxidative and osmotic stress responses in Rhizobium etli. J Bacteriol 191: 4122–4132. |
| [93] |
Reyes-Perez A, Vargas MD, Hernandez M, et al. (2016) Transcriptomic analysis of the process of biofilm formation in Rhizobium etli CFN42. Arch Microbiol 198: 847–860. doi: 10.1007/s00203-016-1241-5
|
| [94] |
Fujishige NA, Kapadia NN, De Hoff PL, et al. (2006) Investigations of rhizobium biofilm formation. FEMS Microbiol Ecol 56: 195–206. doi: 10.1111/j.1574-6941.2005.00044.x
|
| [95] | Glick BR (2012) Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012: 1–15. |
| [96] |
Lucy M, Reed E, Glick BR (2004) Applications of free living plant growth-promoting rhizobacteria. A Van Leeuw J Microb 86: 1–25. doi: 10.1023/B:ANTO.0000024903.10757.6e
|
| [97] |
Stephens JHG, Rask HM (2000) Inoculant production and formulation. Field Crops Res 65: 249–258. doi: 10.1016/S0378-4290(99)00090-8
|
| [98] | Boscari A, Van de Sype G, Le Rudulier D, et al. (2006) Overexpression of BetS, a Sinorhizobium meliloti high-affinity betaine transporter, in bacteroids from Medicago sativa nodules sustains nitrogen fixation during early salt stress adaptation. Mol Plant Microbe In 19: 896–903. |
| [99] |
Elbein AD, Pan YT, Pastuszak I, et al. (2003) New insights on trehalose: a multifunctional molecule. Glycobiology 13: 17R–27R. doi: 10.1093/glycob/cwg047
|
| [100] |
Suarez R, Wong A, Ramirez M, et al. (2008) Improvement of drought tolerance and grain yield in common bean by overexpressing trehalose-6-phosphate synthase in rhizobia. Mol Plant Microbe In 21: 958–966. doi: 10.1094/MPMI-21-7-0958
|
| [101] |
Bari R, Jones JD (2009) Role of plant hormones in plant defence responses. Plant Mol Biol 69: 473–488. doi: 10.1007/s11103-008-9435-0
|
| [102] |
Middleton PH, Jakab J, Penmetsa RV, et al. (2007) An ERF transcription factor in Medicago truncatula that is essential for Nod factor signal transduction. Plant Cell 19: 1221–1234. doi: 10.1105/tpc.106.048264
|
| [103] | Brigido C, Nascimento FX, Duan J, et al. (2013) Expression of an exogenous 1-aminocyclopropane-1-carboxylate deaminase gene in Mesorhizobium spp. reduces the negative effects of salt stress in chickpea. FEMS Microbiol Lett 349: 46–53. |
| [104] |
Kong Z, Glick BR, Duan J, et al. (2015) Effects of 1-aminocyclopropane-1-carboxylate (ACC) deaminase-overproducing Sinorhizobium meliloti on plant growth and copper tolerance of Medicago lupulina. Plant Soil 391: 383–398. doi: 10.1007/s11104-015-2434-4
|
| [105] |
Nascimento FX, Brigido C, Glick BR, et al. (2012) Mesorhizobium ciceri LMS-1 expressing an exogenous 1-aminocyclopropane-1-carboxylate (ACC) deaminase increases its nodulation abilities and chickpea plant resistance to soil constraints. Lett Appl Microbiol 55: 15–21. doi: 10.1111/j.1472-765X.2012.03251.x
|
| [106] |
Nascimento F, Brígido C, Alho L, et al. (2012) Enhanced chickpea growth-promotion ability of a Mesorhizobium strain expressing an exogenous ACC deaminase gene. Plant Soil 353: 221–230. doi: 10.1007/s11104-011-1025-2
|
| [107] |
Bianco C, Defez R (2009) Medicago truncatula improves salt tolerance when nodulated by an indole-3-acetic acid-overproducing Sinorhizobium meliloti strain. J Exp Bot 60: 3097–3107. doi: 10.1093/jxb/erp140
|
| [108] |
Bianco C, Defez R (2010) Improvement of phosphate solubilization and Medicago plant yield by an indole-3-acetic acid-overproducing strain of Sinorhizobium meliloti. Appl Environ Microbiol 76: 4626–4632. doi: 10.1128/AEM.02756-09
|
| [109] |
Shvaleva A, Coba de la Peña T, Rincón A, et al. (2010) Flavodoxin overexpression reduces cadmium-induced damage in alfalfa root nodules. Plant Soil 326: 109–121. doi: 10.1007/s11104-009-9985-1
|
| [110] | Redondo FJ, de la Pena TC, Morcillo CN, et al. (2009) Overexpression of flavodoxin in bacteroids induces changes in antioxidant metabolism leading to delayed senescence and starch accumulation in alfalfa root nodules. Plant Physiol 149: 1166–1178. |
| [111] |
Hill S, Kennedy C, Kavanagh E, et al. (1981) Nitrogen fixation gene (nifL) involved in oxygen regulation of nitrogenase synthesis in K. pneumoniae. Nature 290: 424–426. doi: 10.1038/290424a0
|
| [112] |
Downie JA (2005) Legume haemoglobins: Symbiotic nitrogen fixation needs bloody nodules. Curr Biol 15: R196–R198. doi: 10.1016/j.cub.2005.03.007
|
| [113] |
Talbi C, Sanchez C, Hidalgo-Garcia A, et al. (2012) Enhanced expression of Rhizobium etli cbb(3) oxidase improves drought tolerance of common bean symbiotic nitrogen fixation. J Exp Bot 63: 5035–5043. doi: 10.1093/jxb/ers101
|
| [114] |
Yurgel SN, Soberon M, Sharypova LA, et al. (1998) Isolation of Sinorhizobium meliloti Tn5 mutants with altered cytochrome terminal oxidase expression and improved symbiotic performance. FEMS Microbiol Lett 165: 167–173. doi: 10.1111/j.1574-6968.1998.tb13142.x
|
| [115] | Soberon M, Lopez O, Morera C, et al. (1999) Enhanced nitrogen fixation in a Rhizobium etli ntrC mutant that overproduces the Bradyrhizobium japonicum symbiotic terminal oxidase cbb3. Appl Environ Microbiol 65: 2015–2019. |
| [116] |
Paço A, Brígido C, Alexandre A, et al. (2016) The symbiotic performance of chickpea rhizobia can be improved by additional copies of the clpB chaperone gene. PLoS One 11: e0148221. doi: 10.1371/journal.pone.0148221
|
| [117] |
Alexandre A, Brígido C, Laranjo M, et al. (2009) A survey of chickpea rhizobia diversity in Portugal reveals the predominance of species distinct from Mesorhizobium ciceri and Mesorhizobium mediterraneum. Microb Ecol 58: 930–941. doi: 10.1007/s00248-009-9536-6
|
| [118] | Gibson AH (1987) Evaluation of nitrogen fixation by legumes in the greenhouse and growth chamber, In: Elkan GH, Editor, Symbiotic Nitrogen Fixation Technology, New York: Marcel Dekker, Inc., 321–363. |
| [119] | Beringer JE (1974) R factor transfer in Rhizobium leguminosarum. J Gen Microbiol 84: 188–198. |
| [120] |
Laranjo M, Alexandre A, Oliveira S (2017) Global transcriptional response to salt shock of the plant microsymbiont Mesorhizobium loti MAFF303099. Res Microbiol 168: 55–63. doi: 10.1016/j.resmic.2016.07.006
|
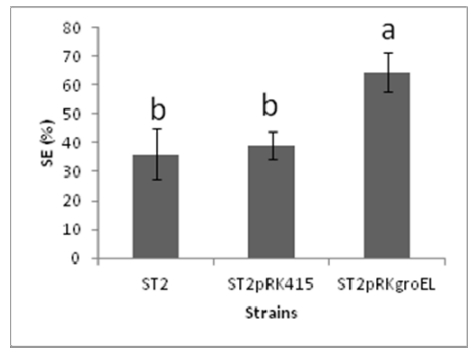
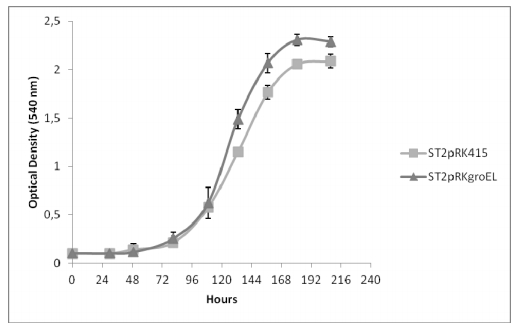
Figures(2)

José Rodrigo da-Silva, Ana Alexandre, Clarisse Brígido, Solange Oliveira. Can stress response genes be used to improve the symbiotic performance of rhizobia?[J]. AIMS Microbiology, 2017, 3(3): 365-382. doi: 10.3934/microbiol.2017.3.365







 DownLoad:
DownLoad: