Citation: Paola Ortiz-Ojeda, Katty Ogata-Gutiérrez, Doris Zúñiga-Dávila. Evaluation of plant growth promoting activity and heavy metal tolerance of psychrotrophic bacteria associated with maca (Lepidium meyenii Walp.) rhizosphere[J]. AIMS Microbiology, 2017, 3(2): 279-292. doi: 10.3934/microbiol.2017.2.279

| [1] | Upadhyay A, Srivastava S (2010) Evaluation of multiple plant growth promoting traits of an isolate of Pseudomonas fluorescens strain Psd. Indian J Exp Biol 48: 601–609. |
| [2] |
Patten CL, Glick BR (2002) Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ Microbiol 68: 3795–3801. doi: 10.1128/AEM.68.8.3795-3801.2002

|
| [3] | Döbereiner J (1992) History and new perspectives of diazotrophs in association with non-leguminous plants. Symbiosis 13: 1–13. |
| [4] | American Public Health Association (1998) Standard methods for examination of water and waste water. Washington DC, USA. |
| [5] | Calvo P, Zúñiga D (2010) Caracterización fisiológica de cepas de Bacillus spp. aisladas de la rizósfera de papa (Solanum tuberosum). Ecol Apl 9: 31–39. |
| [6] |
Ehmann A (1977) The Van Urk-Salkowski reagent a sensitive and specific chromogenic reagent for silica gel thin-layer chromatographic detection and identification of indole derivatives. J Chromatogr 132: 267–276. doi: 10.1016/S0021-9673(00)89300-0
|
| [7] |
Gordon SA, Weber RP (1951) Colorimetric estimation of indolacetic acid. Plant Physiol 26: 192–195. doi: 10.1104/pp.26.1.192
|
| [8] |
Glick BR (1995) The enhancement of plant growth by free-living bacteria. Can J Microbiol 41: 109–117. doi: 10.1139/m95-015
|
| [9] |
Alikhani H, Saleh-Rastin N, Antoun H (2006) Phosphate solubilization activity of rhizobia native to Iranian soils. Plant Soil 287: 35–41. doi: 10.1007/s11104-006-9059-6
|
| [10] |
Nautiyal CS (1999) An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol Lett 170: 265–270. doi: 10.1111/j.1574-6968.1999.tb13383.x
|
| [11] |
Nguyen C, Yan W, Tacon F, et al. (1992) Genetic variability of phosphate solubilizing activity by monocaryotic and dicaryotic mycelia of the ectomycorrhizal fungus Laccaria bicolor (Maire) PD Orton. Plant Soil 143: 193–199. doi: 10.1007/BF00007873
|
| [12] | Ogata K, Zúñiga D (2008) Estudio de la microflora de la rizósfera de Caesalpinia spinosa en la provincia de Huánuco. Zonas Áridas 12: 191–208. |
| [13] | Lalitha MK (2005) Manual on antimicrobial susceptibility testing. Indian Assoc of Med Microbiol 14–16. |
| [14] |
Sabry SA, Ghozlan HA, Abou-Zeid DM (1997) Metal tolerance and antibiotic resistance patterns of a bacterial population isolated from sea water. J Appl Microbiol 82: 245–252. doi: 10.1111/j.1365-2672.1997.tb02858.x
|
| [15] |
Weisburg WG, Barns SM, Pelletier DA, et al. (1991) 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol 173: 697–703. doi: 10.1128/jb.173.2.697-703.1991
|
| [16] |
Larkin MA, Blackshields G, Brown NP, et al. (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23: 2947–2948. doi: 10.1093/bioinformatics/btm404
|
| [17] |
Tamura K, Stecher G, Peterson D, et al. (2013) MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30: 2725–2729. doi: 10.1093/molbev/mst197
|
| [18] | MINAM, Decreto supremo 002-2013 Environmental Quality Standards of agricultural soils, El Peruano, 2012. Available from: http://www.minam.gob.pe/calidadambiental/wp-content/ uploads/sites/22/2013/10/D-S-N-002-2013-MINAM.pdf. |
| [19] | Bric JM, Bostock RM, Silvestone SE (1991) Rapid in situ assay for indolacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl Environ Microbio 57: 535–538. |
| [20] |
Spaepen S, Vanderleyden J, Remans R (2007) Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol Rev 31: 425–448. doi: 10.1111/j.1574-6976.2007.00072.x
|
| [21] | Subramanian P, Kim K, Krishnamoorthy R, et al. (2016) Cold stress tolerance in psychrotolerant soil bacteria and their conferred chilling resistance in tomato (Solanum lycopersicum Mill.) under low temperatures. Plos One 11: 1–17. |
| [22] | Zúñiga-Dávila D, Tolentino J, García M Pérez W, et al. (2011) Characterization of rhizospheric bacteria isolated from maca (Lepidium meyenii Walp.) in the highlands of Junin-Peru, In: Mendez-Vilas A. Editor, Microorganisms in Industry and Environment: From scientific and industrial research to consumer products, Singapore: World Scientific Pub Co, 21–25. |
| [23] | Trivedi P, Pandey A (2007) Low temperature phosphate solubilization and plant growth promotion by psychrotrophic bacteria, isolated from the Himalayan region. Res J Microbiol 2: 452–461. |
| [24] |
Mishra V, Gupta A, Kaur P, et al. (2016) Synergistic effects of Arbuscular mycorrhizal fungi and plant growth promoting rhizobacterial. Int J Phytoremediation 18: 697–703. doi: 10.1080/15226514.2015.1131231
|
| [25] | Kloepper JW, Zablotowicz RM, Tipping EM, et al. (1991) Plant growth promotion mediated by bacterial rhizosphere colonizers, In: Keister DL, Cregan PB, Editors, The rhizosphere and plant growth, Netherlands: Kluwer Academic Publishers, 315–326. |
| [26] | Gul B, Hameed A, Weber DJ, et al. (2016) Assessing seed germination responses of great basin halophytes to various exogenous chemical treatments under saline conditions, In: Sabkha Ecosystems Volume V: The Americas Tasks for Vegetation Science, Switzerland: Springer International Publishing, 85–104. |
| [27] | Acosta JM, Bentivegna DJ, Panigo ES, et al. (2012) Factors affecting seed germination and emergence of Gomphrena perennis. Weed Res 53: 69–75. |
| [28] | Babich H, Stotizky G (1976) Sensitivity of various bacteria, including actinomycetes, and fungi to cadmium and the influence of pH on sensitivity. Appl Environ Microbiol 33: 681–695. |
| [29] |
Franklin NM, Stauber JL, Markich SJ, et al. (2000) pH-dependent toxicity of copper and uranium to a tropical freshwater alga (Chlorella sp.). Aquat Toxicol 48: 275–289. doi: 10.1016/S0166-445X(99)00042-9
|
| [30] |
Sandrin TR, Maier RM (2002). Effect of pH on cadmium toxicity, speciation, and accumulation during naphthalene biodegradation. Environ Toxicol Chem 21: 2075–2079. doi: 10.1002/etc.5620211010
|
| [31] |
Rouch DA, Lee TOB, Morby AP (1995) Understanding cellular responses to toxic agents: a model for mechanisms-choice in bacterial resistance. J Ind Microbiol 14: 132–141. doi: 10.1007/BF01569895
|
| [32] |
Kumar V, Singh S, Bhadrecha P, et al. (2015). Bioremediation of heavy metals by employing resistant microbial isolates from agricultural soil irrigated with industrial waste water. Orient J Chem 31: 357–361. doi: 10.13005/ojc/310142
|
| [33] | Ochiai EI (1987) General principles of biochemistry of the elements, New York: Plenum Press, 381–382. |
| [34] |
Gadd GM (1992) Metals and microorganism: a problem of definition. FEMS Microbiol Lett 100: 197–204. doi: 10.1111/j.1574-6968.1992.tb05703.x
|
| [35] |
Kumar V, Singh S, Singh J, et al. (2015). Potential of plant growth promoting traits by bacteria isolated from heavy metal contaminated soils. Bull Environ Contam Toxicol 94: 807–814. doi: 10.1007/s00128-015-1523-7
|
| [36] | Azario R, Salvarezza S, Ibarra A, et al. (2010) Efecto del cromo hexavalente y trivalente sobre el crecimiento de Escherichia coli ATCC 35218. Información Tecnológica 21: 51–56. |
| [37] | Zahid M, Abbasi MK, Hameed S, et al. (2015) Isolation and identification of indigenous plant growth promoting rhizobacteria from Himalayan region of Kashmir and their effect on improving growth and nutrient contents of maize (Zea mays L.). Front Microbiol 6: 1–10. |
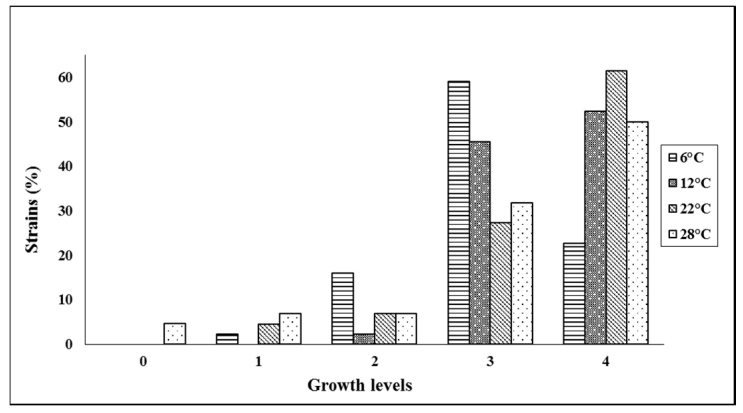
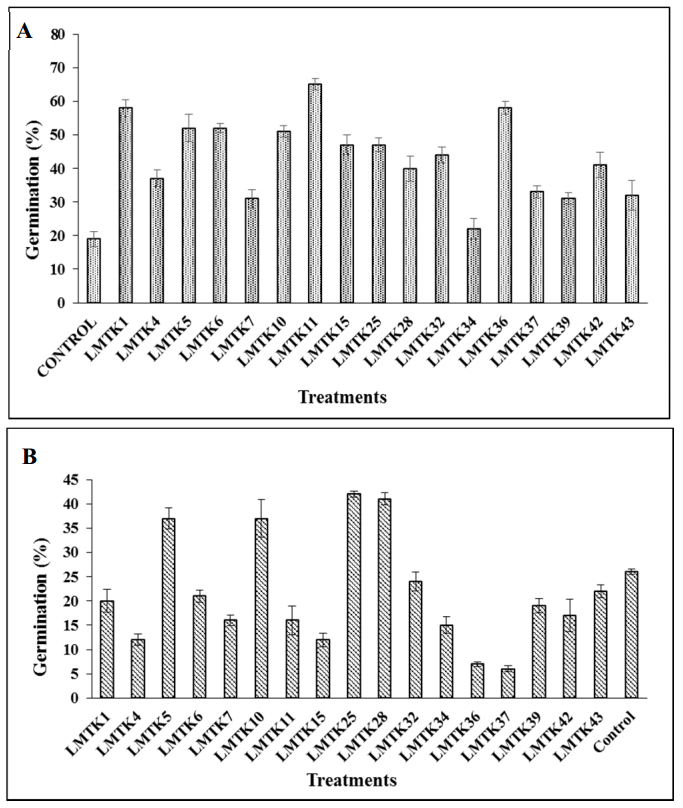
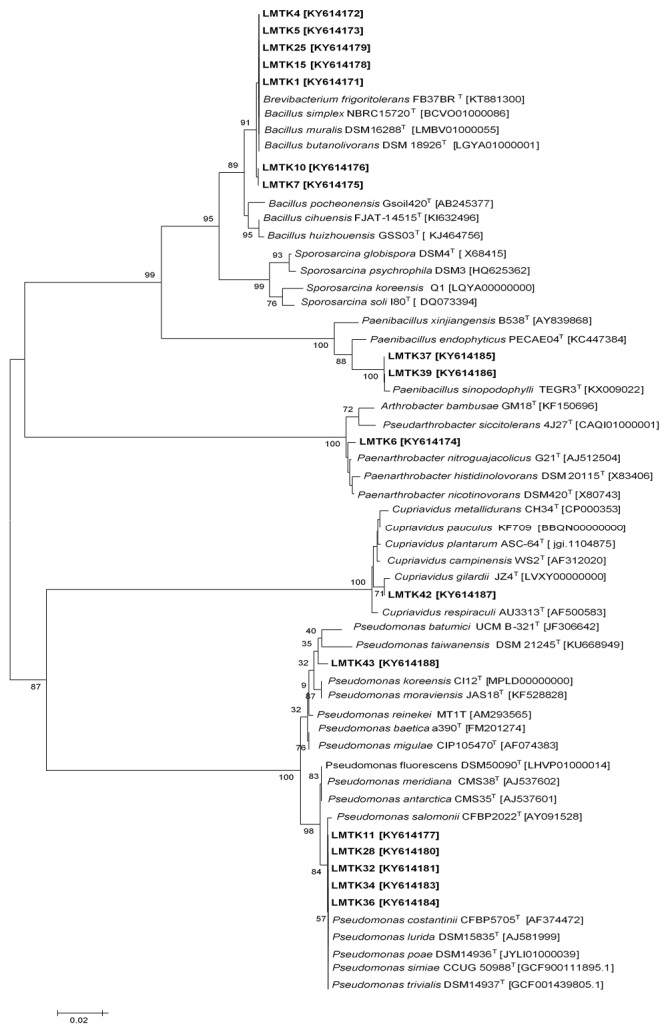
Figures(3) / Tables(4)

Paola Ortiz-Ojeda, Katty Ogata-Gutiérrez, Doris Zúñiga-Dávila. Evaluation of plant growth promoting activity and heavy metal tolerance of psychrotrophic bacteria associated with maca (Lepidium meyenii Walp.) rhizosphere[J]. AIMS Microbiology, 2017, 3(2): 279-292. doi: 10.3934/microbiol.2017.2.279







 DownLoad:
DownLoad: