Citation: Arbaz Sajjad, Wan Zaripah Wan Bakar, Dasmawati Mohamad, Thirumulu Ponnuraj Kannan. Characterization and enhancement of physico-mechanical properties of glass ionomer cement by incorporating a novel nano zirconia silica hydroxyapatite composite synthesized via sol-gel[J]. AIMS Materials Science, 2019, 6(5): 730-747. doi: 10.3934/matersci.2019.5.730

| [1] | Widstrom E, Birn H, Haugejorden O, et al. (1992) Fear of amalgam: dentists' experiences in the Nordic countries. Int Dent J 42: 65–70. |
| [2] |
Pagano S, Chieruzzi M, Balloni S, et al. (2019) Biological, thermal and mechanical characterization of modified glass ionomer cements: the role of nanohydroxyapatite, ciprofloxacin and zinc l-carnosine. Mat Sci Eng C-Mater 94: 76–85. doi: 10.1016/j.msec.2018.09.018

|
| [3] |
Chieruzzi M, Pagano S, Lombardo G, et al. (2018) Effect of nanohydroxyapatite, antibiotic, and mucosal defensive agent on the mechanical and thermal properties of glass ionomer cements for special needs patients. J Mater Res 33: 638–649. doi: 10.1557/jmr.2018.36
|
| [4] | Wilson AD, Kent BE (1971) The glass-ionomer cement, a new translucent dental filling material. J Appl Chem Biotechnol 21: 313. |
| [5] |
Wilson AD, Kent BE (1972) A new translucent cement for dentistry, the glass ionomer cement. Brit Dent J 132: 133–135. doi: 10.1038/sj.bdj.4802810
|
| [6] |
Xie D, Brantley WA, Culbertson BM, et al. (2000) Mechanical properties and microstructures of glass-ionomer cements. Dent Mater 16: 129–138. doi: 10.1016/S0109-5641(99)00093-7
|
| [7] | Anusavice KJ (2003) Mechanical properties of dental materials, Philips Science of Dental Materials, 11 Eds., Philadelphia: Saunders 48–67. |
| [8] |
Bonifacio CC, Kleverlaan CJ, Raggio DP, et al. (2009) Physical-mechanical properties of glass ionomer cements indicated for atraumatic restorative treatment. Aust Dent J 54: 233–237. doi: 10.1111/j.1834-7819.2009.01125.x
|
| [9] | Bonifacio CC, van Amerongen WE, Meschini TG, et al. (2010) Flowable glass ionomer cement as a liner: improving marginal adaptation of atraumatic restorative treatment restorations. J Dent Child (Chic) 77: 12–16. |
| [10] |
Smith DC (1998) Development of glass-ionomer cement systems. Biomaterials 19: 467–478. doi: 10.1016/S0142-9612(97)00126-9
|
| [11] | Smith DC (1968) A new dental cement. Br Dent J 124: 381–384. |
| [12] | Naasan MA, Watson TF (1998) Conventional glass ionomers as posterior restorations. A status report for the American Journal of Dentistry. Am J Dent 11: 36–45. |
| [13] |
Oliva A, Della Ragione F, Salerno A, et al. (1996) Biocompatibility studies on glass ionomer cements by primary cultures of human osteoblasts. Biomaterials 17: 1351–1356. doi: 10.1016/0142-9612(96)88683-2
|
| [14] | Mount GJ (2002) An Atlas of Glass Ionomer Cements: A Clinician's Guide. United Kingdom: Martin Dunitz Ltd. |
| [15] |
Six N, Lasfargues JJ, Goldberg M (2000) In vivo study of the pulp reaction to Fuji IX, a glass ionomer cement. J Dent 28: 413–422. doi: 10.1016/S0300-5712(00)00015-4
|
| [16] | Dimkov A, Nicholson W, Gjorgievska E, et al. (2012) Compressive strength and setting time determination of glass-ionomer cements incorporated with cetylpyridinium chloride and benzalkonium chloride. Prilozi 33: 243–263. |
| [17] | McLean JW, Gasser O (1985) Glass-cermet cements. Quintessence Int 16: 333–343. |
| [18] |
McLean J (1984) Alternatives to amalgam alloys: 1. Br Dent J 157: 432–433. doi: 10.1038/sj.bdj.4805511
|
| [19] |
Sarkar NK (1999) Metal-matrix interface in reinforced glass ionomers. Dent Mater 15: 421–425. doi: 10.1016/S0109-5641(99)00069-X
|
| [20] |
Walls AW, Adamson J, McCabe JF, et al. (1987) The properties of a glass polyalkenoate (ionomer) cement incorporating sintered metallic particles. Dent Mater 3: 113–116. doi: 10.1016/S0109-5641(87)80041-6
|
| [21] | Simmons JJ (1983) The miracle mixture. Glass ionomer and alloy powder. Tex Dent J 100: 6–12. |
| [22] |
Simmons JJ (1990) Silver-alloy powder and glass ionomer cement. J Am Dent Assoc 120: 49–52. doi: 10.14219/jada.archive.1990.0018
|
| [23] |
Nourmohammadi J, Sadrnezhaad SK, Behnamghader A (2010) In vitro bioactivity of novel cured ionomer cement based on iron oxide. Ceram Int 36: 1645–1651. doi: 10.1016/j.ceramint.2010.02.031
|
| [24] |
Kerby R, Bleiholder R (1991) Physical properties of stainless-steel and silver-reinforced glass-ionomer cements. J Dent Res 70: 1358–1361. doi: 10.1177/00220345910700100801
|
| [25] |
Pamir T, Sen BH, Celik A (2005) Mechanical and fluoride release properties of titanium tetrafluoride-added glass-ionomer cement. Dent Mater J 24: 98–103. doi: 10.4012/dmj.24.98
|
| [26] |
Lohbauer U, Walker J, Nikolaenko S, et al. (2003) Reactive fibre reinforced glass ionomer cements. Biomaterials 24: 2901–2907. doi: 10.1016/S0142-9612(03)00130-3
|
| [27] |
Lohbauer U, Frankenberger R, Clare A, et al. (2004) Toughening of dental glass ionomer cements with reactive glass fibres. Biomaterials 25: 5217–5225. doi: 10.1016/j.biomaterials.2003.12.027
|
| [28] | Yamamoto Y (1984) Study on hydroxyapatite-polyacrylic acid composite cement (hydroxyapatite-glass ionomer cement). Shika Zairyo Kikai 3: 787–796. |
| [29] |
Yap AU, Pek YS, Kumar RA, et al. (2002) Experimental studies on a new bioactive material: HAIonomer cements. Biomaterials 23: 955–962. doi: 10.1016/S0142-9612(01)00208-3
|
| [30] |
Moshaverinia M, Borzabadi-Farahani A, Sameni A, et al. (2016) Effects of incorporation of nano-fluorapatite particles on microhardness, fluoride releasing properties, and biocompatibility of a conventional glass ionomer cement (GIC). Dent Mater J 35: 817–821. doi: 10.4012/dmj.2015-437
|
| [31] |
Mitra SB, Wu D, Holmes BN (2003) An application of nanotechnology in advanced dental materials. J Am Dent Assoc 134: 1382–1390. doi: 10.14219/jada.archive.2003.0054
|
| [32] |
Arita K, Lucas ME, Nishino M (2003) The effect of adding hydroxyapatite on the flexural strength of glass ionomer cement. Dent Mater J 22: 126–136. doi: 10.4012/dmj.22.126
|
| [33] |
Arita K, Yamamoto A, Shinonaga Y, et al. (2011) Hydroxyapatite particle characteristics influence the enhancement of the mechanical and chemical properties of conventional restorative glassionomer cement. Dent Mater J 30: 672–683. doi: 10.4012/dmj.2011-029
|
| [34] |
Shiekh RA, Ab Rahman I, Luddin N (2014) Modification of glass ionomer cement by incorporating hydroxyapatite-silica nano-powder composite: sol-gel synthesis and characterization. Ceram Int 40: 3165–3170. doi: 10.1016/j.ceramint.2013.09.127
|
| [35] |
Rahman IA, Masudi SAM, Luddin N, et al. (2014) One-pot synthesis of hydroxyapatite-silica nanopowder composite for hardness enhancement of glass ionomer cement (GIC). B Mater Sci 37: 213–219. doi: 10.1007/s12034-014-0648-3
|
| [36] | Gu YW, Yap AU, Cheang P, et al. (2005) Effects of incorporation of HA/ZrO 2 into glass ionomer cement (GIC). Biomaterials 26: 713–720. |
| [37] |
Gu YW, Yap AUJ, Cheang P, et al. (2005) Developement of zirconia–glass ionomer cement composites. J Non-Cryst Solids 351: 508–514. doi: 10.1016/j.jnoncrysol.2005.01.045
|
| [38] |
Rajabzadeh G, Salehi S, Nemati A, et al. (2014) Enhancing glass ionomer cement features by using the HA/YSZ nanocomposite: a feed forward neural network modelling. J Mech Behav Biomed Mater 29: 317–327. doi: 10.1016/j.jmbbm.2013.07.025
|
| [39] |
Silva VV, Lameiras FS, Lobato ZI (2002) Biological reactivity of zirconia-hydroxyapatite composites. J Biomed Mater Res 63: 583–590. doi: 10.1002/jbm.10308
|
| [40] |
Chou BY, Chang E (2002) Plasma-sprayed hydroxyapatite coating on titanium alloy with ZrO2 second phase and ZrO2 intermediate layer. Surf Coat Tech 153: 84–92. doi: 10.1016/S0257-8972(01)01532-8
|
| [41] |
Rahman IA, Ghazali NAM, Bakar WZW, et al. (2017) Modification of glass ionomer cement by incorporating nanozirconia-hydroxyapatite-silica nano-powder composite by the one-pot technique for hardness and aesthetics improvement. Ceram Int 43: 13247–13253. doi: 10.1016/j.ceramint.2017.07.022
|
| [42] |
Panda R, Hsieh M, Chung R, et al. (2003) FTIR, XRD, SEM and solid state NMR investigations of carbonate-containing hydroxyapatite nano-particles synthesized by hydroxide-gel technique. J Phys Chem Solids 64: 193–199. doi: 10.1016/S0022-3697(02)00257-3
|
| [43] |
Moheet IA, Luddin N, Ab Rahman I, et al. (2018) Evaluation of mechanical properties and bond strength of nano-hydroxyapatite-silica added glass ionomer cement. Ceram Inte 44: 9899–9906. doi: 10.1016/j.ceramint.2018.03.010
|
| [44] |
Perez Cdos R, Hirata RJ, da Silva AH, et al. (2009) Effect of a glaze/composite sealant on the 3-D surface roughness of esthetic restorative materials. Oper Dent 34: 674–680. doi: 10.2341/08-014-L
|
| [45] |
Takei T, Kato K, Meguro A, et al. (1999) Infrared spectra of geminal and novel triple hydroxyl groups on silica surface. Colloid Surface A 150: 77–84. doi: 10.1016/S0927-7757(98)00813-9
|
| [46] | Rahman I, Masudi SaM, Luddin N, et al. (2013) One-pot synthesis of hydroxyapatite-silica nano-powder composite for hardness enhancement of glass ionomer cement (GIC). B Mater Sci 37: 213–219. |
| [47] |
Zhan Z, Zeng HC (1999) A catalyst-free approach for sol-gel synthesis of highly mixed ZrO2–SiO2 oxides. J Non-Crys Solids 243: 26–38. doi: 10.1016/S0022-3093(98)00810-2
|
| [48] |
Fernando MS, de Silva RM, de Silva KMN (2015) Synthesis, characterization, and application of nano hydroxyapatite and nanocomposite of hydroxyapatite with granular activated carbon for the removal of Pb 2+ from aqueous solutions. Appl Surf Sci 351: 95–103. doi: 10.1016/j.apsusc.2015.05.092
|
| [49] |
Wang A, Liu D, Yin H, et al. (2007) Size-controlled synthesis of hydroxyapatite nanorods by chemical precipitation in the presence of organic modifiers. Mat Sci Eng C-Mater 27: 865–869. doi: 10.1016/j.msec.2006.10.001
|
| [50] | Singh AK, Nakate UT (2014) Microwave synthesis, characterization, and photoluminescence properties of nanocrystalline zirconia. Sci World J 2014: 349457. |
| [51] | Ranjbar M, Yousefi M, Lahooti M, et al. (2012) Preparation and characterization of tetragonal zirconium oxide nanocrystals from isophthalic acid-zirconium(IV) nanocomposite as a new precursor. Int J Nanosci Nanotechno 8: 191–196. |
| [52] |
Xiao Y, Li D, Fan H, et al. (2007) Preparation of nano-HA/PLA composite by modified-PLA for controlling the growth of HA crystals. Mater Lett 61: 59–62. doi: 10.1016/j.matlet.2006.04.005
|
| [53] |
Nejati E, Mirzadeh H, Zandi M (2008) Synthesis and characterization of nano-hydroxyapatite rods/poly(L-lactide acid) composite scaffolds for bone tissue engineering. Compos Part A-Appl S 39: 1589–1596. doi: 10.1016/j.compositesa.2008.05.018
|
| [54] |
Stoch A, Jastrzębski W, Brożek A, et al. (2000) FTIR absorption-reflection study of biomimetic growth of phosphates on titanium implants. J Mol Struct 555: 375–382. doi: 10.1016/S0022-2860(00)00623-2
|
| [55] |
de Bruijn JD, Bovell YP, van Blitterswijk CA (1994) Structural arrangements at the interface between plasma sprayed calcium phosphates and bone. Biomaterials 15: 543–550. doi: 10.1016/0142-9612(94)90021-3
|
| [56] |
Jafarzadeh M, Rahman IA, Sipaut CS (2009) Synthesis of silica nanoparticles by modified sol-gel process: the effect of mixing modes of the reactants and drying techniques. J Sol-Gel Sci Techn 50: 328–336. doi: 10.1007/s10971-009-1958-6
|
| [57] |
Rahman IA, Vejayakumaran P, Sipaut CS, et al. (2007) An optimized sol-gel synthesis of stable primary equivalent silica particles. Colloid Surface A 294: 102–110. doi: 10.1016/j.colsurfa.2006.08.001
|
| [58] |
Moheet IA, Luddin N, Ab Rahman I, et al. (2018) Evaluation of mechanical properties and bond strength of nano-hydroxyapatite-silica added glass ionomer cement. Ceram Int 44: 9899–9906. doi: 10.1016/j.ceramint.2018.03.010
|
| [59] |
Gu YW, Yap AUJ, Cheang PHN, et al. (2005) Zirconia-glass ionomer cement-a potential substitute for miracle mix. Scripta Mater 52: 113–116. doi: 10.1016/j.scriptamat.2004.09.019
|
| [60] | Anusavice KJ (2003) Mechanical properties of dental materials, In: Anusavice KJ, Shen C, Rawls HR, Philips Science of Dental Materials, 11 Eds., New York: Saunders, 48–67. |
| [61] |
Takaki P, Vieira M, Bommarito S (2014) Maximum bite force analysis in different age groups. Int Arch Otorhinolaryngol 18: 272–276. doi: 10.1055/s-0034-1374647
|
| [62] | Rios D, Honório HM, Araújo PAd, et al. (2002) Wear and superficial roughness of glass ionomer cements used as sealants, after simulated toothbrushing. Pesqui Odontol Bras16: 343–348. |
| [63] |
Yip HK, Lam WT, Smales RJ (1999) Fluoride release, weight loss and erosive wear of modern aesthetic restoratives. Br Dent J 187: 265–270. doi: 10.1038/sj.bdj.4800256
|
| [64] | Yip HK, Lam WT, Smales RJ (1999) Surface roughness and weight loss of esthetic restorative materials related to fluoride release and uptake. J Clin Pediatr Dent 23: 321–326. |
| [65] | Yip KH, Peng D, Smales RJ (2001) Effects of APF gel on the physical structure of compomers and glass ionomer cements. Oper Dent 26: 231–238. |
| [66] | Mount GJ (1995) Some physical and biological properties of glass ionomer cement. Int Dent J 45: 135–140. |
| [67] | Lucas ME, Arita K, Nishino M (2003) Toughness, bonding and fluoride-release properties |
| [68] | of hydroxyapatite-added glass ionomer cement. Biomaterials 24: 3787–3794. |
| [69] |
68. Cales B, Stefani Y, Lilley E (1994) Long-term in vivo and in vivo aging of a zirconia ceramic used in orthopaedy. J Biomed Mater Res 28: 619–624. doi: 10.1002/jbm.820280512
|
| [70] | 69. Uo M, Sjoren G, Sundh A, et al. Cytotoxicity and bonding property of dental ceramics. Dent Mater 19: 487–492. |
| [71] |
70. Moshaverinia A, Ansari S, Moshaverinia M, et al. (2008) Effects of incorporation of hydroxyapatite and fluoroapatite nanobioceramic into conventional galss ionomer cements. Acta Biomater 4: 432–440. doi: 10.1016/j.actbio.2007.07.011
|
| [72] |
71. Piconi C, Maccauro G (1999) Zirconia as a ceramic biomaterial. Biomaterials 20: 1–25. doi: 10.1016/S0142-9612(98)00010-6
|
| [73] | 72. Aoki H (1991) Science and Medical Applications of Hydroxyapatite, Tokyo: JAAS. |
| [74] |
73. Shinonaga Y, Arita K, Nishimura T, et al. (2015) Effects of porous-hydroxyapatite incorporated into glass-ionomer sealants. Dent Mater J 34: 196–202. doi: 10.4012/dmj.2014-195
|
| [75] |
74. Lopes CMCF, Galvan J, Chibinski ACR, et al. (2018) Fluoride release and surface roughness of a new glass ionomer cement: glass carbomer. Rev Odontol UNESP 47: 1–6. doi: 10.1590/1807-2577.06717
|
| [76] | 75. Bala O, Arisu HD, Yikilgan I, et al. (2012) Evaluation of surface roughness and hardness of different glass ionomer cements. Eur J Dent 6: 79–86. |
| [77] |
76. Bollen CM, Lambrechts P, Quirynen M (1997) Comparison of surface roughness of oral hard materials to the threshold surface roughness for bacterial plaque retention: a review of the literature. Dent Mater 13: 258–269. doi: 10.1016/S0109-5641(97)80038-3
|
| [78] |
77. Warren DP, Colescott TD, Henson HA, et al. (2002) Effects of four prophylaxis pastes on surface roughness of a composite, a hybrid ionomer, and a compomer restorative material. J Esthet Restor Den 14: 245–251. doi: 10.1111/j.1708-8240.2002.tb00170.x
|
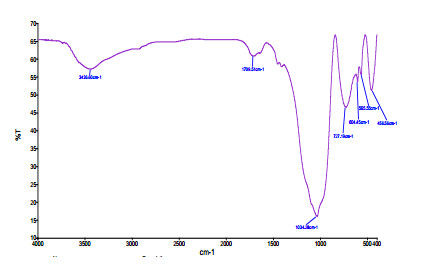
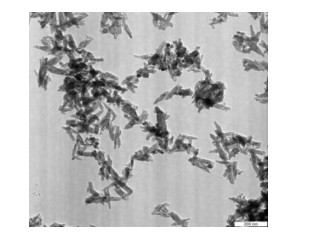
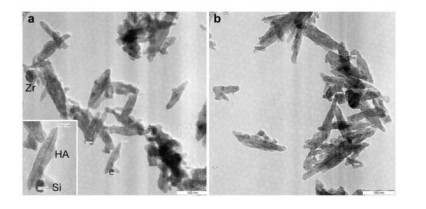

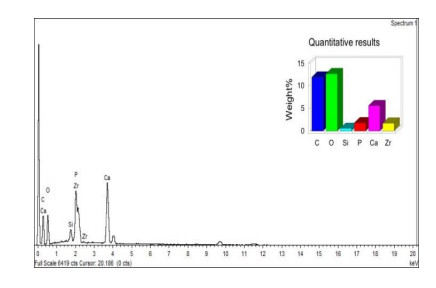

Figures(6) / Tables(3)

Arbaz Sajjad, Wan Zaripah Wan Bakar, Dasmawati Mohamad, Thirumulu Ponnuraj Kannan. Characterization and enhancement of physico-mechanical properties of glass ionomer cement by incorporating a novel nano zirconia silica hydroxyapatite composite synthesized via sol-gel[J]. AIMS Materials Science, 2019, 6(5): 730-747. doi: 10.3934/matersci.2019.5.730







 DownLoad:
DownLoad: