Citation: Laura J. Weiser, Erik E. Santiso. Molecular modeling studies of peptoid polymers[J]. AIMS Materials Science, 2017, 4(5): 1029-1051. doi: 10.3934/matersci.2017.5.1029

| [1] |
Sun J, Zuckermann RN (2013) Peptoid Polymers: A Highly Designable Bioinspired Material. ACS Nano 7: 4715–4732. doi: 10.1021/nn4015714

|
| [2] | Seo J, Lee BC, Zuckermann RN (2011) Peptoids: Synthesis, Characterization, and Nanostructures. Compr Biomater 2: 53–76. |
| [3] |
Chongsiriwatana NP, Patch JA, Czyzewski AM, et al. (2008) Peptoids that mimic the structure, function, and mechanism of helical antimicrobial peptides. Proc Natl Acad Sci USA 105: 2794–2799. doi: 10.1073/pnas.0708254105
|
| [4] |
Vollrath SBL, Fürniss D, Schepers U, et al. (2013) Amphiphilic peptoid transporters-synthesis and evaluation. Org Biomol Chem 11: 8197–8201. doi: 10.1039/c3ob41139g
|
| [5] |
Li N, Zhu F, Gao F, et al. (2010) Blockade of CD28 by a synthetical peptoid inhibits T-cell proliferation and attenuates graft-versus-host disease. Cell Mol Immunol 7: 133–142. doi: 10.1038/cmi.2009.120
|
| [6] |
Dohm MT, Kapoor R, Barron AE (2011) Peptoids: Bio-Inspired Polymers as Potential Pharmaceuticals. Curr Pharm Design 17: 2732–2747. doi: 10.2174/138161211797416066
|
| [7] |
Statz AR, Park JP, Chongsiriwatana NP, et al. (2008) Surface-immobilised antimicrobial peptoids. Biofouling 24: 439–448. doi: 10.1080/08927010802331829
|
| [8] |
Seurynck SL, Patch JA, Barron AE (2005) Simple, helical peptoid analogs of lung surfactant protein B. Chem Biol 12: 77–88. doi: 10.1016/j.chembiol.2004.10.014
|
| [9] |
Maayan G, Ward MD, Kirshenbaum K (2009) Folded biomimetic oligomers for enantioselective catalysis. Proc Natl Acad Sci USA 106: 13679–13684. doi: 10.1073/pnas.0903187106
|
| [10] | Gellman SH (1998) Foldamers: A Manifesto. Accounts Chem Res 31: 173–180. |
| [11] |
Armand P, Kirshenbaum K, Falicov A, et al. (1997) Chiral N-substituted glycines can form stable helical conformations. Fold Design 2: 369–375. doi: 10.1016/S1359-0278(97)00051-5
|
| [12] |
Shah NH, Butterfoss GL, Nguyen K (2008) Oligo(N-aryl glycines): A New Twist on Structured Peptoids. J Am Chem Soc 130: 16622–16632. doi: 10.1021/ja804580n
|
| [13] |
Huang K, Wu CW, Sanborn TJ, et al. (2006) A threaded loop conformation adopted by a family of peptoid nonamers. J Am Chem Soc 128: 1733–1738. doi: 10.1021/ja0574318
|
| [14] |
Crapster JA, Guzei IA, Blackwell HE (2013) A Peptoid Ribbon Secondary Structure. Angew Chem Int Ed 52: 5079–5084. doi: 10.1002/anie.201208630
|
| [15] |
Mannige RV, Haxton TK, Proulx C, et al. (2015) Peptoid nanosheets exhibit a new secondary-structure motif. Nature 526: 415–420. doi: 10.1038/nature15363
|
| [16] |
Hebert ML, Shah DS, Blake P, et al. (2013) Tunable peptoid microspheres: effects of side chain chemistry and sequence. Org Biomol Chem 11: 4459–4464. doi: 10.1039/c3ob40561c
|
| [17] |
Murnen HK, Rosales AM, Jaworski JN, et al. (2010) Hierarchical Self-Assembly of a Biomimetic Diblock Copolypeptoid into Homochiral Superhelices. J Am Chem Soc 132: 16112–16119. doi: 10.1021/ja106340f
|
| [18] |
Sanii B, Kudirka R, Cho A, et al. (2011) Shaken, Not Stirred: Collapsing a Peptoid Monolayer To Produce Free-Floating, Stable Nanosheets. J Am Chem Soc 133: 20808–20815. doi: 10.1021/ja206199d
|
| [19] |
Dill KA, MacCallum JL (2012) The Protein-Folding Problem, 50 Years On. Science 338: 1042–1046. doi: 10.1126/science.1219021
|
| [20] |
Gorske BC, Blackwell HE (2006) Tuning peptoid secondary structure with pentafluoroaromatic functionality: A new design paradigm for the construction of discretely folded peptoid structures. J Am Chem Soc 128: 14378–14387. doi: 10.1021/ja065248o
|
| [21] |
Stringer JR, Crapster JA, Guzei IA, et al. (2010) Construction of Peptoids with All Trans-Amide Backbones and Peptoid Reverse Turns via the Tactical Incorporation of N-Aryl Side Chains Capable of Hydrogen Bonding. J Org Chem 75: 6068–6078. doi: 10.1021/jo101075a
|
| [22] |
Gorske BC, Nelson RC, Bowden ZS, et al. (2013) "Bridged" n→π* Interactions Can Stabilize Peptoid Helices. J Org Chem 78: 11172–11183. doi: 10.1021/jo4014113
|
| [23] |
Wu CW, Kirshenbaum K, Sanborn TJ, et al. (2003) Structural and spectroscopic studies of peptoid oligomers with alpha-chiral aliphatic side chains. J Am Chem Soc 125: 13525–13530. doi: 10.1021/ja037540r
|
| [24] |
Kirshenbaum K, Barron AE, Goldsmith RA, et al. (1998) Sequence-specific polypeptoids: A diverse family of heteropolymers with stable secondary structure. Proc Natl Acad Sci USA 95: 4303–4308. doi: 10.1073/pnas.95.8.4303
|
| [25] |
Armand P, Kirshenbaum K, Goldsmith RA, et al. (1998) NMR determination of the major solution conformation of a peptoid pentamer with chiral side chains. Proc Natl Acad Sci USA 95: 4309–4314. doi: 10.1073/pnas.95.8.4309
|
| [26] | Dill KA (1990) Dominant forces in protein folding. Biochemistry 29: 31. |
| [27] |
Dill KA, Ozkan SB, Shell MS, et al. (2008) The protein folding problem. Annu Rev Biophys 37: 289–316. doi: 10.1146/annurev.biophys.37.092707.153558
|
| [28] |
Sali A, Blundell TL (1993) Comparative protein modeling by satisfaction of spatial restraints. J Mol Biol 234: 779–815. doi: 10.1006/jmbi.1993.1626
|
| [29] |
Shen MY, Sali A (2006) Statistical potential for assessment and prediction of protein structures. Protein Sci 15: 2507–2524. doi: 10.1110/ps.062416606
|
| [30] |
Rohl CA, Strauss CEM, Misura KMS, et al. (2004) Protein structure prediction using Rosetta. Method Enzymol 383: 66–93. doi: 10.1016/S0076-6879(04)83004-0
|
| [31] | Leach AR (2001) Molecular modelling : principles and applications, England: Pearson/Prentice Hall. |
| [32] | Shell MS (2016) Coarse-Graining with the Relative Entropy, In: Rice SA, Dinner AR, Advances in Chemical Physics, Malden: Wiley-Blackwell, 395–441. |
| [33] |
Mackerell AD, Feig M, Brooks CL (2004) Extending the treatment of backbone energetics in protein force fields: Limitations of gas-phase quantum mechanics in reproducing protein conformational distributions in molecular dynamics simulations. J Comput Chem 25: 1400–1415. doi: 10.1002/jcc.20065
|
| [34] |
Feigel M (1983) Rotation barriers of amides in the gas phase. J Phys Chem 87: 3054–3058. doi: 10.1021/j100239a019
|
| [35] |
Sui Q, Borchardt D, Rabenstein DL (2007) Kinetics and equilibria of cis/trans isomerization of backbone amide bonds in peptoids. J Am Chem Soc 129: 12042–12048. doi: 10.1021/ja0740925
|
| [36] |
Duffy EM, Severance DL, Jorgensen WL (1992) Solvent effects on the barrier to isomerization for a tertiary amide from ab initio and Monte Carlo calculations. J Am Chem Soc 114: 7535–7542. doi: 10.1021/ja00045a029
|
| [37] |
Torrie GM, Valleau JP (1977) Nonphysical sampling distributions in Monte Carlo free-energy estimation: Umbrella sampling. J Comput Phys 23: 187–199. doi: 10.1016/0021-9991(77)90121-8
|
| [38] |
Sugita Y, Okamoto Y (1999) Replica-exchange molecular dynamics method for protein folding. Chem Phys Lett 314: 141–151. doi: 10.1016/S0009-2614(99)01123-9
|
| [39] |
Stringer JR, Crapster JA, Guzei IA, et al. (2011) Extraordinarily Robust Polyproline Type I Peptoid Helices Generated via the Incorporation of alpha-Chiral Aromatic N-1-Naphthylethyl Side Chains. J Am Chem Soc 133: 15559–15567. doi: 10.1021/ja204755p
|
| [40] |
Ramachandran GN, Ramakrishnan C, Sasisekharan V (1963) Stereochemistry of polypeptide chain configurations. J Mol Biol 7: 95–99. doi: 10.1016/S0022-2836(63)80023-6
|
| [41] |
Butterfoss GL, Renfrew PD, Kuhlman B, et al. (2009) A Preliminary Survey of the Peptoid Folding Landscape. J Am Chem Soc 131: 16798–16807. doi: 10.1021/ja905267k
|
| [42] |
Mohle K, Hofmann HJ (1996) Peptides and peptoids—A systematic structure comparison. J Mol Model 2: 307–311. doi: 10.1007/s0089460020307
|
| [43] | Miertuš S, Scrocco E, Tomasi J (1981) Electrostatic interaction of a solute with a continuum. A direct utilizaion of AB initio molecular potentials for the prevision of solvent effects. Chem Phys 55: 117–129. |
| [44] | Pascual-Ahuir JL, Silla E, Tomasi J, et al. (1987) Electrostatic interaction of a solute with a continuum. Improved description of the cavity and of the surface cavity bound charge distribution. J Comput Chem 8: 778–787. |
| [45] |
Parker BF, Knight AS, Vukovic S, et al. (2016) A Peptoid-Based Combinatorial and Computational Approach to Developing Ligands for Uranyl Sequestration from Seawater. Ind Eng Chem Res 55: 4187–4194. doi: 10.1021/acs.iecr.5b03500
|
| [46] |
Cancès E, Mennucci B, Tomasi J (1997) A new integral equation formalism for the polarizable continuum model: Theoretical background and applications to isotropic and anisotropic dielectrics. J Chem Phys 107: 3032–3041. doi: 10.1063/1.474659
|
| [47] |
Cornell W, Cieplek P, Bayly CI, et al. (1995) A Second Generation Force-Field for the Simulation of Proteins, Nucleic-Acids, and Organic-Molecules. J Am Chem Soc 117: 5179–5197. doi: 10.1021/ja00124a002
|
| [48] |
Hawkins GD, Cramer CJ, Truhlar DG (1998) Universal Quantum Mechanical Model for Solvation Free Energies Based on Gas-Phase Geometries. J Phys Chem B 102: 3257–3271. doi: 10.1021/jp973306+
|
| [49] |
Bradley EK, Kerr JM, Richter LS, et al. (1997) NMR Structural Characterization of Oligo-N-Substituted Glycine Lead Compounds from a Combinatorial Library. Mol Divers 3: 1–15. doi: 10.1023/A:1009698309407
|
| [50] | Mann G, Yun RH, Nyland L, et al. (2002) The Sigma MD Program and a Generic Interface Applicable to Multi-Functional Programs with Complex, Hierarchical Command Structure, In: Schlick T, Gan HH, Computational Methods for Macromolecules: Challenges and Applications, Springer, Berlin, Heidelberg, 129–145. |
| [51] |
Hermans J, Berendsen HJC, Van Gunsteren WF, et al. (1984) A consistent empirical potential for water–protein interactions. Biopolymers 23: 1513–1518. doi: 10.1002/bip.360230807
|
| [52] |
Butterfoss GL, Yoo B, Jaworski JN (2012) De novo structure prediction and experimental characterization of folded peptoid oligomers. Proc Natl Acad Sci USA 109: 14320–14325. doi: 10.1073/pnas.1209945109
|
| [53] |
Wang JM, Wolf RM, Caldwell JW, et al. (2004) Development and testing of a general amber force field. J Comput Chem 25: 1157–1174. doi: 10.1002/jcc.20035
|
| [54] |
Onufriev A, Bashford D, Case DA (2004) Exploring protein native states and large-scale conformational changes with a modified generalized born model. Proteins 55: 383–394. doi: 10.1002/prot.20033
|
| [55] |
MacKerell AD, Bashford D, Bellott M, et al. (1998) All-atom empirical potential for molecular modeling and dynamics studies of proteins. J Phys Chem B 102: 3586–3616. doi: 10.1021/jp973084f
|
| [56] |
Case DA, Cheatham TE, Darden T, et al. (2005) The Amber biomolecular simulation programs. J Comput Chem 26: 1668–1688. doi: 10.1002/jcc.20290
|
| [57] |
Jorgensen WL, Maxwell DS, TiradoRives J (1996) Development and testing of the OPLS all-atom force field on conformational energetics and properties of organic liquids. J Am Chem Soc 118: 11225–11236. doi: 10.1021/ja9621760
|
| [58] |
Moehle K, Hofmann HJ (1996) Peptides and peptoids—A quantum chemical structure comparison. Biopolymers 38: 781–790. doi: 10.1002/(SICI)1097-0282(199606)38:6<781::AID-BIP9>3.0.CO;2-N
|
| [59] |
Jorgensen W, Chandrasekhar J, Madura J, et al. (1983) Comparison of Simple Potential Functions for Simulating Liquid Water. J Chem Phys 79: 926–935. doi: 10.1063/1.445869
|
| [60] |
Tobias DJ, Brooks CL (1988) Molecular dynamics with internal coordinate constraints. J Chem Phys 89: 5115–5127. doi: 10.1063/1.455654
|
| [61] |
Wang J, Wang W, Kollman PA, et al. (2006) Automatic atom type and bond type perception in molecular mechanical calculations. J Mol Graph Model 25: 247–260. doi: 10.1016/j.jmgm.2005.12.005
|
| [62] | Jakalian A, Jack DB, Bayly CI (2002) Fast, efficient generation of high-quality atomic charges. AM1-BCC model: II. Parameterization and validation. J Comput Chem 23: 1623–1641. |
| [63] |
Mukherjee S, Zhou G, Michel C, et al. (2015) Insights into Peptoid Helix Folding Cooperativity from an Improved Backbone Potential. J Phys Chem B 119: 15407–15417. doi: 10.1021/acs.jpcb.5b09625
|
| [64] |
Lifson S, Roig A (1961) On the Theory of Helix-Coil Transition in Polypeptides. J Chem Phys 34: 1963–1974. doi: 10.1063/1.1731802
|
| [65] |
Mirijanian DT, Mannige RV, Zuckermann RN, et al. (2014) Development and use of an atomistic CHARMM-based forcefield for peptoid simulation. J Comput Chem 35: 360–370. doi: 10.1002/jcc.23478
|
| [66] |
Jordan PA, Bishwajit P, Butterfoss GL, et al. (2011) Oligo(N-alkoxy glycines): trans substantiating peptoid conformations. J Pept Sci 96: 617–626. doi: 10.1002/bip.21675
|
| [67] |
Nam KT, Shelby SA, Cho PH, et al. (2010) Free-floating ultrathin two-dimensional crystals from sequence-specific peptoid polymers. Nat Mater 9: 454–460. doi: 10.1038/nmat2742
|
| [68] |
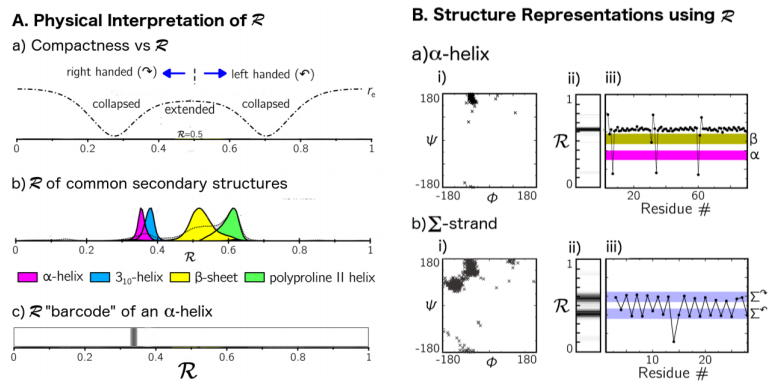
Mannige RV, Kundu J, Whitelam S (2016) The Ramachandran Number: An Order Parameter for Protein Geometry. PLoS One 11: e0160023. doi: 10.1371/journal.pone.0160023
|
| [69] |
Reith D, Putz M, Muller-Plathe F (2003) Deriving effective mesoscale potentials from atomistic simulations. J Comput Chem 24: 1624–1636. doi: 10.1002/jcc.10307
|
| [70] |
Izvekov S, Voth GA (2005) Multiscale coarse graining of liquid-state systems. J Chem Phys 123: 134105. doi: 10.1063/1.2038787
|
| [71] |
Shell MS (2008) The relative entropy is fundamental to multiscale and inverse thermodynamic problems. J Chem Phys 129: 144108. doi: 10.1063/1.2992060
|
| [72] |
Haxton TK, Mannige RV, Zuckermann RN, et al. (2015) Modeling Sequence-Specific Polymers Using Anisotropic Coarse-Grained Sites Allows Quantitative Comparison with Experiment. J Chem Theory Comput 11: 303–315. doi: 10.1021/ct5010559
|
| [73] |
Sanii B, Haxton TK, Olivier GK, et al. (2014) Structure-Determining Step in the Hierarchical Assembly of Peptoid Nanosheets. ACS Nano 8: 11674–11684. doi: 10.1021/nn505007u
|
| [74] |
Haxton TK, Zuckermann RN, Whitelam S (2016) Implicit-Solvent Coarse-Grained Simulation with a Fluctuating Interface Reveals a Molecular Mechanism for Peptoid Monolayer Buckling. J Chem Theory Comput 12: 345–352. doi: 10.1021/acs.jctc.5b00910
|
| [75] | Drew K, Renfrew PD, Butterfoss GL (2013) Adding Diverse Noncanonical Backbones to Rosetta: Enabling Peptidomimetic Design. PLoS One 8: e67051. |
| [76] |
Kaufmann KW, Lemmon GH, DeLuca SL, et al. (2010) Practically Useful: What the Rosetta Protein Modeling Suite Can Do for You. Biochemistry 49: 2987–2998. doi: 10.1021/bi902153g
|
| [77] | Renfrew PD, Craven TW, Butterfoss GL, et al. (2013) A Rotamer Library to Enable Modeling and Design of Peptoid Foldamers. J Am Chem Soc 136: 8772–8782. |
| [78] |
Laio A, Gervasio FL (2008) Metadynamics: a method to simulate rare events and reconstruct the free energy in biophysics, chemistry and material science. Rep Prog Phys 71: 126601. doi: 10.1088/0034-4885/71/12/126601
|
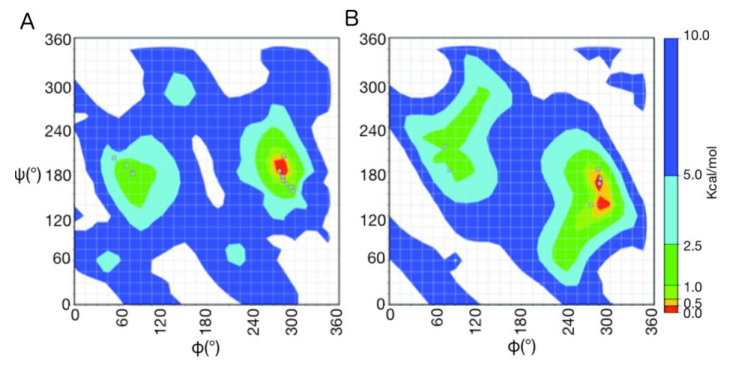
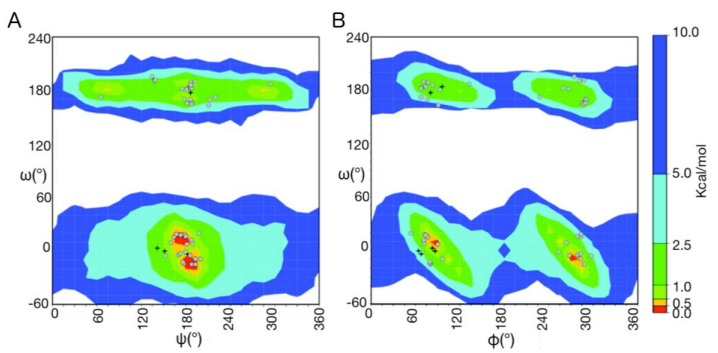
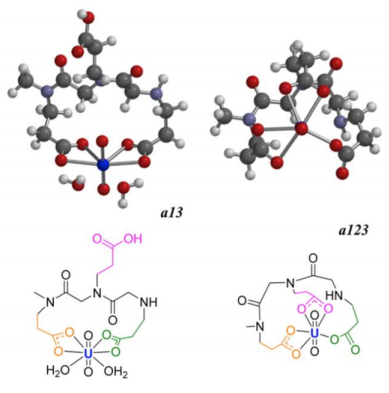
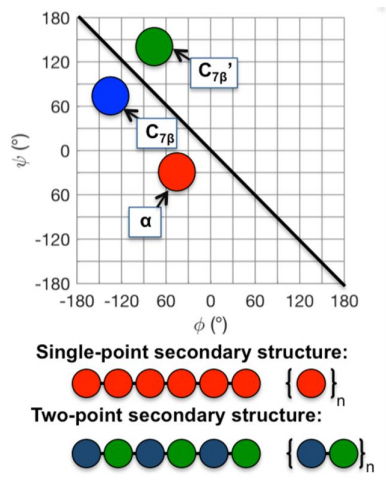
Figures(11)

Laura J. Weiser, Erik E. Santiso. Molecular modeling studies of peptoid polymers[J]. AIMS Materials Science, 2017, 4(5): 1029-1051. doi: 10.3934/matersci.2017.5.1029







 DownLoad:
DownLoad: