Citation: Uichi Akiba, Jun-ichi Anzai. Cyclodextrin-containing layer-by-layer films and microcapsules: Synthesis and applications[J]. AIMS Materials Science, 2017, 4(4): 832-846. doi: 10.3934/matersci.2017.4.832

| [1] |
Gentile P, Carmagnola I, Nardo T, et al. (2015) Layer-by-layer assembly for biomedical applications in the last decade. Nanotechnology 26: 422001. doi: 10.1088/0957-4484/26/42/422001

|
| [2] |
Barsan MM, Brett CMA (2016) Recent advances in layer-by-layer strategies for biosensors incorporating metal nanoparticles. TrAC-Trend Anal Chem 79: 286–296. doi: 10.1016/j.trac.2015.11.019
|
| [3] |
Richardson JJ, Cui J, Björnmalm M, et al. (2016) Innovation in layer-by-layer assembly. Chem Rev 116: 14828–14867. doi: 10.1021/acs.chemrev.6b00627
|
| [4] |
Xuan M, Zhao J, Shao J, et al. (2017) Recent progress in layer-by-layer assembled biogenic capsules and their applications. J Colloid Interf Sci 487: 107–117. doi: 10.1016/j.jcis.2016.10.018
|
| [5] |
Polomska A, Leroux J, Brambilla D (2017) Layer-by-layer coating of solid drug cores: A versatile method to improve stability, control release and tune surface properties. Macromol Biosci 17: 1600228. doi: 10.1002/mabi.201600228
|
| [6] |
Anzai J, Kobayashi Y, Suzuki Y, et al. (1998) Enzyme sensors prepared by layer-by-layer deposition of enzymes on a platinum electrode through avidin-biotin interaction. Sensor Actuat B-Chem 52: 3–9. doi: 10.1016/S0925-4005(98)00248-2
|
| [7] |
Shi H, Yang Y, Huang JD, et al. (2006) Amperometric choline biosensors prepared by layer-by-layer deposition of choline oxidase on the Prussian blue-modified platinum electrode. Talanta 70: 852–858. doi: 10.1016/j.talanta.2006.02.012
|
| [8] |
Wang B, Anzai J (2007) Redox reactions of ferricyanide ions in layer-by-layer deposited polysaccharide films: A significant effect of the type of polycation in the films. Langmuir 23: 7378–7384. doi: 10.1021/la070119m
|
| [9] |
Crouzier T, Boudou T, Picart C (2010) Polysaccharide-based polyelectrolyte multilayers. Curr Opin Colloid In 15: 417–426. doi: 10.1016/j.cocis.2010.05.007
|
| [10] |
Sato H, Anzai J (2006) Preparation of layer-by-layer thin films composed of DNA and ferrocene-bearing poly(amine)s and their redox properties. Biomacromolecules 7: 2072–2076. doi: 10.1021/bm060222a
|
| [11] |
Jewell CM, Lynn DM (2008) Multilayered polyelectrolyte assemblies as platforms for the delivery of DNA and other nucleic acid-based therapeutics. Adv Drug Deliver Rev 60: 979–999. doi: 10.1016/j.addr.2008.02.010
|
| [12] |
Sukhishvili SA, Granick S (2002) Layered, erasable polymer multilayers formed by hydrogen-bonded sequential self-assembly. Macromolecules 35: 301–310. doi: 10.1021/ma011346c
|
| [13] |
Pavlukhina S, Sukhishvili S (2011) Polymer assemblies for controlled delivery of bioactive molecules from surfaces. Adv Drug Deliver Rev 63: 822–836. doi: 10.1016/j.addr.2011.03.017
|
| [14] |
Rao SV, Anderson KW, Bachas LG (1999) Controlled layer-by-layer immobilization of horseradish peroxidase. Biotechnol Bioeng 65: 389–396. doi: 10.1002/(SICI)1097-0290(19991120)65:4<389::AID-BIT3>3.0.CO;2-V
|
| [15] |
Sato K, Kodama D, Naka Y, et al. (2006) Electrochemically induced disintegration of layer-by-layer-assembled thin films composed of 2-iminobiotin-labeled poly(ethyleneimine) and avidin. Biomacromolecules 7: 3302–3305. doi: 10.1021/bm060819q
|
| [16] |
Hoshi T, Akase S, Anzai J (2002) Preparation of multilayer thin films containing avidin through sugar-lectin interactions and their binding properties. Langmuir 18: 7024–7028. doi: 10.1021/la0203685
|
| [17] |
Donath E, Sukhorukov GB, Caruso F, et al. (1998) Novel hollow polymer shells by colloid-templated assembly of polyelectrolytes. Angew Chem Int Ed 37: 2201–2205. doi: 10.1002/(SICI)1521-3773(19980904)37:16<2201::AID-ANIE2201>3.0.CO;2-E
|
| [18] | Antipov AA, Sukhorukov GB, Leporatti S, et al. (2002) Polyelectrolyte multilayer capsule permeability control. Colloid Surface A 198–200: 535–541. |
| [19] |
De Mercato LL, Ferraro MM, Baldassarre F, et al. (2014) Biological applications of LBL multilayer capsules: From drug delivery to sensing. Adv Colloid Interfac 207: 139–154. doi: 10.1016/j.cis.2014.02.014
|
| [20] |
Liu L, Son M, Park H, et al. (2014) Efficacy of CNT-bound polyelecby spray-assisted layer-by-layer (LbL) technique on water purification. RSC Adv 4: 32858–32865. doi: 10.1039/C4RA05272B
|
| [21] |
Kaner P, Johnson DJ, Seker E, et al. (2015) Layer-by-layer surface modification of polyethersulfone membranes using polyelectrolytes and AgCl/TiO2 xerogels. J Membrane Sci 493: 807–819. doi: 10.1016/j.memsci.2015.05.048
|
| [22] |
Sato K, Takahashi S, Anzai J (2012) Layer-by-layer thin films and microcapsules for biosensors and controlled release. Anal Sci 28: 929–938. doi: 10.2116/analsci.28.929
|
| [23] |
Wu B, Hou S, Huang L, et al. (2008) Oriented immobilization of immunoglobulin G onto the cuvette of the resonant mirror biosensor through layer-by-layer assembly of multilayer films. Mater Sci Eng C 28: 1065–1069. doi: 10.1016/j.msec.2007.04.035
|
| [24] |
Yoshida K, Hasebe Y, Takahashi S, et al. (2014) Layer-by-layer deposited nano- and micro-assemblies for insulin delivery: A review. Mater Sci Eng C 34: 384–392. doi: 10.1016/j.msec.2013.09.045
|
| [25] |
Hashide R, Yoshida K, Hasebe Y, et al. (2014) Poly(lactic acid) microparticles coated with insulin-containing layer-by-layer films and their pH-dependent insulin release. J Nanosci Nanotechno 14: 3100–3105. doi: 10.1166/jnn.2014.8562
|
| [26] |
Zhu Y, Xuan H, Ren J, et al. (2015) Self-healing multilayer polyelectrolyte composite film with chitosan and poly(acrylic acid). Soft Matt 11: 8452–8459. doi: 10.1039/C5SM01463H
|
| [27] |
Zhu Y, Yin T, Ren J, et al. (2016) Self-healing polyelectrolyte multilayer composite film with microcapsules. RSC Adv 6: 12100–12106. doi: 10.1039/C5RA22821B
|
| [28] |
Kurkov SV, Loftsson T (2013) Cyclodextrins. Int J Pharmaceut 453: 167–180. doi: 10.1016/j.ijpharm.2012.06.055
|
| [29] | Suzuki I, Obata K, Anzai J, et al. (2000) Crown ether-tethered cyclodextrins: superiority of the secondary-hydroxy side modification in binding tryptophan. J Chem Soc Perkin Trans 2: 1705–1710. |
| [30] |
Zhu X, Wang H, Chen Q, et al. (2007) Preparation and characterization of inclusion complex of iprodione and b-cyclodextrin to improve fungicidal activity. J Agr Food Chem 55: 3535–3539. doi: 10.1021/jf070197f
|
| [31] |
Takahashi K, Morimoto S, Nakamura H, et al. (2011) Improvement of pharmaceutical potential of all-trans retinoic acid with hydroxypropyl-b-cyclodextrin. J Incl Phenom Macro 70: 389–396. doi: 10.1007/s10847-010-9857-6
|
| [32] |
Moussa Z, Hmadeh M, Abiad MG, et al. (2016) Encapsulation of curcumine in cyclodextrin-metal organic frameworks: Dissociation of loaded Cd-MOFs enhances stability of curcumin. Food Chem 212: 485–494. doi: 10.1016/j.foodchem.2016.06.013
|
| [33] | Čepo DV, Jug M, Rajkovič MG, et al. (2017) Formulation of a nutraceutical derived from carob: b-cyclodextrin encapsulation of antioxidants from carob pod. J Food Nutr Res 56: 48–60. |
| [34] |
Suzuki I, Murakami K, Anzai J (2001) Fabrication of surface-confined ferrocene-appended cyclodextrins on gold electrode. Mater Sci Eng C 17: 149–154. doi: 10.1016/S0928-4931(01)00325-3
|
| [35] |
Arima H, Motoyama K, Higashi T (2017) Potential use of cyclodextrins as drug carriers and active pharmaceutical ingredients. Chem Pharm Bull 65: 341–348. doi: 10.1248/cpb.c16-00779
|
| [36] |
Seki T, Abe K, Egawa Y, et al. (2016) A pseudopolyrotaxane for glucose-responsive insulin release: The effect of binding ability and spatial arrangement of phenylboronic acid group. Mol Pharmaceut 13: 3807–3815. doi: 10.1021/acs.molpharmaceut.6b00599
|
| [37] |
Kojima Y, Okano T, Seki T, et al. (2017) Polyol-responsive pseudopolyrotaxanes based on phenylboronic acid-modified polyethylene glycol and cyclodextrins. J Incl Phenom Macro 87: 295–303. doi: 10.1007/s10847-017-0699-3
|
| [38] |
Yang X, Johnson S, Shi J, et al. (1997) Polyelectrolyte and molecular host ion self-assembly to multilayer thin films: An approach to thin chemical sensors. Sensor Actuat B-Chem 45: 87–92. doi: 10.1016/S0925-4005(97)00274-8
|
| [39] |
Sato K, Suzuki I, Anzai J (2003) Preparation of polyelectrolyte-layered assemblies containing cyclodextrin and their binding properties. Langmuir 19: 7406–7412. doi: 10.1021/la030163b
|
| [40] |
Sato K, Suzuki I, Anzai J (2004) Layered assemblies composed of sulfonated cyclodextrin and poly(allylamine). Colloid Polym Sci 282: 287–290. doi: 10.1007/s00396-003-0911-4
|
| [41] |
Suzki I, Sato K, Koga M, et al. (2003) Polyelectrolyte layered assemblies containing azobenzene-modified polymer and anionic cyclodextrins. Mater Sci Eng C 23: 579–583. doi: 10.1016/S0928-4931(03)00050-X
|
| [42] |
Zhi J, Tian X, Zhao W, et al. (2008) Self-assembled film based on carboxymethyl-b-cyclodextrin and diazoresin and its binding properties for methylene blue. J Colloid Interf Sci 319: 270–276. doi: 10.1016/j.jcis.2007.11.031
|
| [43] |
Yang SY, Hoonor R, Jin H, et al. (2013) Synthesis and characterization of cationic and anionic cyclodextrin oligomaers and their use in layer-by-layer film formation. B Korean Chem Soc 34: 2016–2022. doi: 10.5012/bkcs.2013.34.7.2016
|
| [44] |
Dam HH, Caruso F (2013) Formation and degradation of layer-by-layer-assembled polyelectrolyte polyrotaxane capsules. Langmuir 29: 7203–7208. doi: 10.1021/la304580x
|
| [45] | Suzuki I, Egawa Y, Mizukawa Y, et al. (2002) Construction of positively-charged layered assemblies assisted by cyclodextrin complexation. Chem Commun 164–166. |
| [46] | Heyden AV, Wilczewski M, Labbé P, et al. (2006) Multilayer films based on host-guest interactions between biocompatible polymers. Chem Commun 3220–3222. |
| [47] |
Kaftan O, Tumbiolo S, Dubreuil F, et al. (2011) Probing multivalent host-guest interactions between modified polymer layers by direct force measurement. J Phys ChemB 115: 7726–7735. doi: 10.1021/jp110939c
|
| [48] |
Dubacheva GV, Dumy P, Auzély R, et al. (2010) Unlimited growth of host-guest multilayer films based on functionalized neutral polymers. Soft Matt 6: 3747–3750. doi: 10.1039/c0sm00324g
|
| [49] |
Zhu M, Aryal GH, Zhang N, et al. (2015) Host-guest interactions derived multilayer perylene diimide thin film constructed on a scaffolding porphyrin monolayer. Langmuir 31: 578–586. doi: 10.1021/la504297w
|
| [50] |
Huang J, Yang Y, Shi H, et al. (2006) Multi-walled carbon nanotubes-based glucose biosensor prepared by layer-by-layer technique. Mater Sci Eng C 26: 113–117. doi: 10.1016/j.msec.2005.06.002
|
| [51] |
Zhang S, Vlémincq C, Wong DR, et al. (2016) Nanopapers of layer-by-layer nanotubes. J Mater Chem B 4: 7651–7661. doi: 10.1039/C6TB02737G
|
| [52] | Trejo NK, Frey M (2015) A comparative study on electrosprayed, layer-by-layer, and chemically grafted nanomembranes loaded with iron oxide nanoparticles. J Appl Polym Sci 132: 42657. |
| [53] |
Xiong F, Chen C, Liu S (2016) Preparation of chitosan/polystyrene sulfonate multilayered composite metal nanoparticles and its application. J Nanosci Nanotechno 16: 6027–6031. doi: 10.1166/jnn.2016.12111
|
| [54] |
Crespo-Biel O, Dordi B, Reinhoudt DN, et al. (2005) Supramolecular layer-by-layer assembly: Alternating adsorption of guest- and host-functionalized molecules and particles using multivalent supramolecular interactions. J Am Chem Soc 127: 7594–7600. doi: 10.1021/ja051093t
|
| [55] |
Crespo-Biel O, Dordi B, Maury P, et al. (2006) Patterned, hybrid, multilayer nanostructures based on multivalent supramolecular interactions. Chem Mater 18: 2545–2551. doi: 10.1021/cm052796c
|
| [56] |
Li G, Dong Z, Zhu Y, et al. (2016) Dual-responsive colloidal microcapsules based on host-guest interaction on solid templates. J Colloid Interf Sci 475: 196–202. doi: 10.1016/j.jcis.2016.04.050
|
| [57] |
Zhang Y, Yang RH, Liu F, et al. (2004) Fluorescent sensor for imidazole derivatives based on monomer-dimer equilibrium of a zinc porphyrin complex in a polymeric film. Anal Chem 76: 7336–7345. doi: 10.1021/ac049477+
|
| [58] |
Khan R, Dhayal M (2009) Chitosan/polyaniline hybrid conducting biopolymer base impedimetric immunosensor to detect Ochratoxin-A. Biosens Bioelectron 24: 1700–1705. doi: 10.1016/j.bios.2008.08.046
|
| [59] |
Kurita R, Hirata Y, Yabuki S, et al. (2008) Surface modification of thin polyion complex film for surface plasmon resonance immunosensor. Sensor Actuat B-Chem 130: 320–325. doi: 10.1016/j.snb.2007.08.007
|
| [60] | Anzai J, Ueno A, Osa T (1984) High and rapid response in photo-induced potential changes across a poly(vinyl chloride)/spirobenzopyran membrane. J Chem Soc Chem Commun 688–689. |
| [61] |
Kirsch J, Siltanen C, Zhou Q, et al. (2013) Biosensor technology: recent advances in threat agent detection and medicine. Chem Soc Rev 42: 8733–8768. doi: 10.1039/c3cs60141b
|
| [62] |
Takahashi S, Anzai J (2013) Recent progress in ferrocene-modified thin films and nanoparticles for biosensors. Materials 6: 5742–5762. doi: 10.3390/ma6125742
|
| [63] |
Rao PV, Gan SH (2015) Recent advances in nanotechnology-based diagnosis and treatments of diabetes. Curr Drug Metab 16: 371–375. doi: 10.2174/1389200215666141125120215
|
| [64] |
Zhang W, Shi Y, Zheng S, et al. (2017) Preparation of photoactive multilayer films with high photocurrent response and detection of thrombin. J Electroanal Chem 784: 85–90. doi: 10.1016/j.jelechem.2016.10.061
|
| [65] |
Yang Y, Yang X, Liu Y, et al. (2005) Optical sensor for lithocholic acid based on multilayered assemblies from polyelectrolyte and cyclodextrin. J Photoch Photobio A 171: 137–144. doi: 10.1016/j.jphotochem.2004.10.006
|
| [66] |
Damos FS, Luz RCS, Tanaka AA, et al. (2010) Development of an electroactive layer-by-layer assembly based on host-guest supramolecular interactions. J Electroanal Chem 639: 36–42. doi: 10.1016/j.jelechem.2009.11.008
|
| [67] |
Damos FS, Luz RCS, Tanaka AA, et al. (2010) Dissolved oxygen amperometric sensor based on layer-by-layer assembly using host-guest supramolecular interactions. Anal Chim Acta 664: 144–150. doi: 10.1016/j.aca.2010.02.011
|
| [68] |
Chen W, Luo G, Qiu W, et al. (2017) Mesoporous silica-based versatile theranostic nanoplatform constructed by layer-by-layer assembly for excellent photodynamic/chemo therapy. Biomaterials 117: 54–65. doi: 10.1016/j.biomaterials.2016.11.057
|
| [69] |
Nag S, Duarte L, Bertrand E, et al. (2014) Ultrasensitive QRS made by supramolecular assembly of functionalized cyclodextrins and graphene for the detection of lung cancer VOC biomarkers. J Mater Chem B 2: 6571–6579. doi: 10.1039/C4TB01041H
|
| [70] |
Duarte L, Nag S, Castro M, et al. (2016) Chemical sensors based on new polyamides biobased on (Z) octadec-9-enedioic acid and b-cyclodextrin. Macromol Chem Phys 217: 1620–1628. doi: 10.1002/macp.201600102
|
| [71] |
Li J, Liu D, Tian L, et al. (2009) Electrochemical investigation of myoglobin in layer-by-layer films assembled with sulfonated-b-cyclodextrin. Electroanalysis 21: 2653–2659. doi: 10.1002/elan.200900308
|
| [72] |
Camacho C, Matías JC, Cao R, et al. (2008) Hydrogen peroxide biosensor with a supramolecular layer-by-layer design. Langmuir 24: 7654–7657. doi: 10.1021/la800242a
|
| [73] |
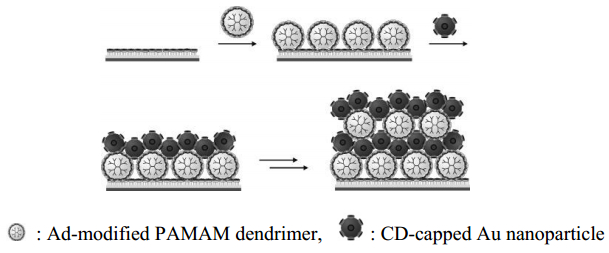
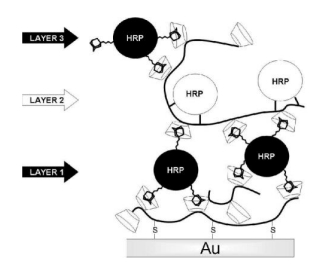
Villalonga R, Díez P, Gamella M, et al. (2012) Layer-by-layer supramolecular architecture of cyclodextrin-modified PAMAM dendrimers and adamantane-modified peroxidase on gold surface for electrochemical biosensing. Electrochim Acta 76: 249–255. doi: 10.1016/j.electacta.2012.05.025
|
| [74] |
Zhu X, Loh X (2015) Layer-by-layer assemblies for antibacterial applications. Biomater Sci 3: 1505–1518. doi: 10.1039/C5BM00307E
|
| [75] |
Wei T, Zhan W, Cao L, et al. (2016) Multifunctional and regenerable antibacterial surfaces fabricated by a universal strategy. ACS Appl Mater Interfaces 8: 30048–30057. doi: 10.1021/acsami.6b11187
|
| [76] |
Qu Y, Wei T, Zhan W, et al. (2017) A reusable supramolecular platform for the specific capture and release of proteins and bacteria. J Mater Chem B 5: 444–453. doi: 10.1039/C6TB02821G
|
| [77] |
Xu G, Pranantyo D, Xu L, et al. (2016) Antifouling, antimicrobial, and antibiocorrosion multilayer coatings assembled by layer-by-layer deposition involving host-guest interaction. Ind Eng Chem Res 55: 10906–10915. doi: 10.1021/acs.iecr.6b02190
|
| [78] |
Wang Z, Feng Z, Gao C (2008) Stepwise assembly of the same polyelectrolytes using host-guest interaction to obtain microcapsules with multiresponsive properties. Chem Mater 20: 4194–4199. doi: 10.1021/cm8003358
|
| [79] |
Li C, Luo G, Wang H, et al. (2011) Host-guest assembly of pH-responsive degradable microcapsules with controlled drug release behavior. J Phys Chem C 115: 17651–17659. doi: 10.1021/jp203940s
|
| [80] |
Luo G, Xu X, Zhang J, et al. (2012) Encapsulation of an adamantane-doxorubicin prodrug in pH-responsive polysaccharide capsules for controlled release. ACS Appl Mater Interfaces 4: 5317–5324. doi: 10.1021/am301258a
|
| [81] |
Xiao W, Chen W, Zhang J, et al. (2011) Design of a photoswitchable hollow microcapsular drug delivery system by using a supramolecular drug-loading approach. J Phys Chem B 115: 13796–13802. doi: 10.1021/jp208692c
|
| [82] | Lin H, Xiao W, Qin S, et al. (2014) Switch on/off microcapsules for controlled photosensitive drug release in a 'release-cease-recommence' mode. Polym Chem 5: 4437–4440. |
| [83] |
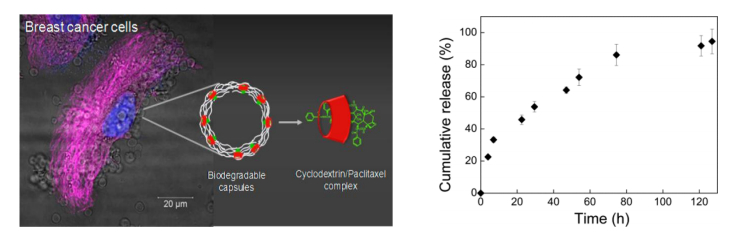
Jing J, Szarpak-Jankowska A, Guillot R, et al. (2013) Cyclodextrin/paclitaxel complex in biodegradable capsules for breast cancer treatment. Chem Mater 25: 3867–3873. doi: 10.1021/cm4019925
|
| [84] | Tejashri G, Amrita B, Darshana J (2013) Cyclodextrin based nanosponges for pharmaceutical use: A review. Acta Pharmaceut 63: 335–358. |
| [85] |
Cho E, Jung S (2015) Supramolecular complexation of carbohydrates for the bioavailability enhancement of poorly soluble drugs. Molecules 20: 19620–19646. doi: 10.3390/molecules201019620
|
| [86] |
Antoniuk I, Amiel C (2016) Cyclodextrin-mediated hierarchical self-assembly and its potential in drug delivery applications. J Pharm Sci 105: 2570–2588. doi: 10.1016/j.xphs.2016.05.010
|
| [87] |
Kurapati R, Raichur AM (2013) Composite cyclodextrin-calcium carbonate porous microparticles and modified multilayer capsules: novel carriers for encapsulation of hydrophobic drugs. J Mater Chem B 1: 3175–3184. doi: 10.1039/c3tb20192a
|
| [88] |
Martin A, Tabary N, Leclercq L, et al. (2013) Multilayered textile coating based on a b-cyclodextrin polyelectrolyte for the controlled release of drugs. Carbohyd Polym 93: 718–730. doi: 10.1016/j.carbpol.2012.12.055
|
| [89] |
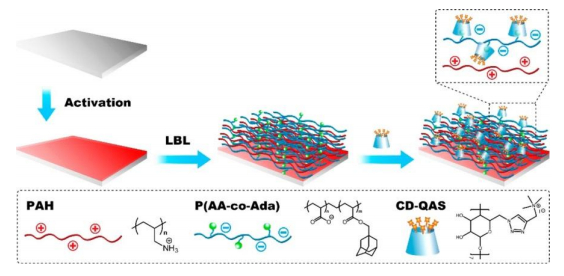
Junthip J, Tabary N, Chai F, et al. (2016) Layer-by-layer coating of textile with two oppositely charged cyclodextrin polyelectrolytes for extended drug delivery. J Biomed Mater Res A 104: 1408–1424. doi: 10.1002/jbm.a.35674
|
| [90] |
Gómez-Galván F, Pérez-Álvarez L, Matas J, et al. (2016) Preparation and characterization of soluble branched ionic b-cyclodextrin and their inclusion complexes with triclosan. Carbohyd Polym 142: 149–157. doi: 10.1016/j.carbpol.2016.01.046
|
| [91] |
Pérez-Álvarez L, Matas J, Gómez-Galván F, et al. (2017) Branched and ionic b-cyclodextrin multilayer assembling onto polyacrylonitrile membranes for removal and controlled release of triclosan. Carbohyd Polym 156: 143–151. doi: 10.1016/j.carbpol.2016.09.020
|
| [92] |
Hashide R, Yoshida K, Hasebe Y, et al. (2012) Insulin-containing layer-by-layer films deposited on poly(lactic acid) microbeads for pH-controlled release of insulin. Colloid Surface B 89: 242–247. doi: 10.1016/j.colsurfb.2011.09.023
|
| [93] |
Masood F (2016) Polymeric nanoparticles for targeted drug delivery system for cancer therapy. Mater Sci Eng C 60: 569–578. doi: 10.1016/j.msec.2015.11.067
|
| [94] |
Fagui AE, Wintgens V, Gaillet C, et al. (2014) Layer-by-layer coated PLA nanoparticles with oppositely charged b-cyclodextrin polymer for controlled delivery of lipophilic molecules. Macromol Chem Phys 215: 555–565. doi: 10.1002/macp.201300693
|
| [95] |
Xuan H, Ren J, Zhang J, et al. (2017) Novel highly-flexible, acid-resistant and self-healing host-guest transparent multilayer films. Appl Surf Sci 411: 303–314. doi: 10.1016/j.apsusc.2017.03.129
|
| [96] |
Xuan H, Ren J, Wang X, et al. (2017) Flame-retardant, non-irritating and self-healing multilayer films with double-network structure. Compos Sci Technol 145: 15–23. doi: 10.1016/j.compscitech.2017.03.038
|
Figures(8)

Uichi Akiba, Jun-ichi Anzai. Cyclodextrin-containing layer-by-layer films and microcapsules: Synthesis and applications[J]. AIMS Materials Science, 2017, 4(4): 832-846. doi: 10.3934/matersci.2017.4.832







 DownLoad:
DownLoad: