Citation: Hiroki Nara, Keisuke Morita, Tokihiko Yokoshima, Daikichi Mukoyama, Toshiyuki Momma, Tetsuya Osaka. Electrochemical impedance spectroscopy analysis with a symmetric cell for LiCoO2 cathode degradation correlated with Co dissolution[J]. AIMS Materials Science, 2016, 3(2): 448-459. doi: 10.3934/matersci.2016.2.448

| [1] | Nishi Y (2001) Lithium ion secondary batteries; past 10 years and the future. J Power Sources 100: 101–106. |
| [2] |
Etacheri V, Marom R, Elazari R, et al. (2011) Challenges in the development of advanced Li-ion batteries: a review. Energy Environ Sci 4: 3243–3262. doi: 10.1039/c1ee01598b

|
| [3] |
Dunn B, Kamath H, Tarascon J-M (2011) Electrical energy storage for the grid: a battery of choices. Science 334: 928–935. doi: 10.1126/science.1212741
|
| [4] |
Yang Z, Zhang J, Kintner-Meyer MCW, et al. (2011) Electrochemical Energy Storage for Green Grid. Chem Rev 111: 3577–3613. doi: 10.1021/cr100290v
|
| [5] |
Broussely M, Biensan P, Bonhomme F, et al. (2005) Main aging mechanisms in Li ion batteries. J Power Sources 146: 90–96. doi: 10.1016/j.jpowsour.2005.03.172
|
| [6] | Vetter J, Novák P, Wagner MR, et al. (2005) Ageing mechanisms in lithium-ion batteries. J Power Sources 147: 269–281. |
| [7] |
Smart MC, Ratnakumar BV (2011) Effects of Electrolyte Composition on Lithium Plating in Lithium-Ion Cells. J Electrochem Soc 158: A379–A389. doi: 10.1149/1.3544439
|
| [8] | Aurbach D, Markovsky B, Salitra G, et al. (2007) Talyossef, M. Koltypin, et al., Review on electrode–electrolyte solution interactions, related to cathode materials for Li-ion batteries. J Power Sources 165: 491–499. |
| [9] |
Aurbach D, Markovsky B, Rodkin A, et al. (2002) On the capacity fading of LiCoO2 intercalation electrodes. Electrochimica Acta 47: 4291–4306. doi: 10.1016/S0013-4686(02)00417-6
|
| [10] |
Takami N, Satoh A, Hara M, et al. (1995) Structural and Kinetic Characterization of Lithium Intercalation into Carbon Anodes for Secondary Lithium Batteries. J Electrochem Soc 142: 371–379. doi: 10.1149/1.2044017
|
| [11] |
Aurbach D, Levi MD, Levi E, et al. (1998) Common Electroanalytical Behavior of Li Intercalation Processes into Graphite and Transition Metal Oxides. J Electrochem Soc 145: 3024–3034. doi: 10.1149/1.1838758
|
| [12] |
Markovsky B, Levi MD, Aurbach D (1998) The basic electroanalytical behavior of practical graphite–lithium intercalation electrodes. Electrochimica Acta 43: 2287–2304. doi: 10.1016/S0013-4686(97)10172-4
|
| [13] | Barsoukov E, Kim JH, Yoon CO, et al. (1999) Kinetics of lithium intercalation into carbon anodes: in situ impedance investigation of thickness and potential dependence. Solid State Ionics. |
| [14] |
Zhang D, Haran BS, Durairajan A, et al. (2000) Studies on capacity fade of lithium-ion batteries. J Power Sources 91: 122–129. doi: 10.1016/S0378-7753(00)00469-9
|
| [15] |
Nagasubramanian G (2000) Two- and three-electrode impedance studies on 18650 Li-ion cells. J Power Sources 87: 226–229. doi: 10.1016/S0378-7753(99)00469-3
|
| [16] |
Li J, Murphy E, Winnick J, et al. (2001) Studies on the cycle life of commercial lithium ion batteries during rapid charge–discharge cycling. J Power Sources 102: 294–301. doi: 10.1016/S0378-7753(01)00821-7
|
| [17] |
Seki S, Kihira N, Mita Y, et al. (2011) AC Impedance Study of High-Power Lithium-Ion Secondary Batteries—Effect of Battery Size. J Electrochem Soc 158: A163–A166. doi: 10.1149/1.3525277
|
| [18] |
Mukoyama D, Momma T, Nara H, et al. (2012) Electrochemical Impedance Analysis on Degradation of Commercially Available Lithium Ion Battery during Charge–Discharge Cycling. Chem Lett 41: 444–446. doi: 10.1246/cl.2012.444
|
| [19] | Osaka T, Momma T, Mukoyama D, et al. (2012) Proposal of novel equivalent circuit for electrochemical impedance analysis of commercially available lithium ion battery. J Power Sources 205: 483–486. |
| [20] |
Hang T, Mukoyama D, Nara H, et al. (2013) Electrochemical impedance spectroscopy analysis for lithium-ion battery using Li4Ti5O12 anode. J Power Sources 222: 442–447. doi: 10.1016/j.jpowsour.2012.09.010
|
| [21] |
Illig J, Ender M, Weber A, et al. (2015) Modeling graphite anodes with serial and transmission line models. J Power Sources 282: 335–347. doi: 10.1016/j.jpowsour.2015.02.038
|
| [22] |
Hoshi Y, Narita Y, Honda K, et al. (2015) Optimization of reference electrode position in a three-electrode cell for impedance measurements in lithium-ion rechargeable battery by finite element method. J Power Sources 288: 168–175. doi: 10.1016/j.jpowsour.2015.04.065
|
| [23] | Osaka T, Mukoyama D, Nara H (2015) Review—Development of Diagnostic Process for Commercially Available Batteries, Especially Lithium Ion Battery, by Electrochemical Impedance Spectroscopy. J Electrochem Soc 162: A2529–A2537. |
| [24] |
Osaka T, Nara H, Mukoyama D, et al. (2013) New Analysis of Electrochemical Impedance Spectroscopy for Lithium-ion Batteries. J Electrochem Sci Tech 4: 157–162. doi: 10.5229/JECST.2013.4.4.157
|
| [25] |
Mendoza-Hernandez OS, Ishikawa H, Nishikawa Y, et al. (2014) State of Charge Dependency of Graphitized-Carbon-Based Reactions in a Lithium-ion Secondary Cell Studied by Electrochemical Impedance Spectroscopy. Electrochimica Acta 131: 168–173. doi: 10.1016/j.electacta.2014.01.057
|
| [26] | Bünzli C, Kaiser H, Novák P (2015) Important Aspects for Reliable Electrochemical Impedance Spectroscopy Measurements of Li-Ion Battery Electrodes. J Electrochem Soc 162: A218–A222. |
| [27] | Yokoshima T, Nara H, Mukoyama D, et al. (2011) Performance of Fine Reference Electrode in Thin Laminated Li-Ion Cell, in: Meet. Abstr, The Electrochemical Society, pp. 1451–1451. |
| [28] | Jansen AN, Dees DW, Abraham DP, et al. ((2007)) Low-temperature study of lithium-ion cells using a LiySn micro-reference electrode. J Power Sources 174: 373–379. |
| [29] | La Mantia F, Wessells CD, Deshazer HD, et al. (2013) Reliable reference electrodes for lithium-ion batteries. Electrochem Commun 31: 141–144. |
| [30] |
Gómez-Cámer JL, Novák P (2013) Electrochemical impedance spectroscopy: Understanding the role of the reference electrode. Electrochem Commun 34: 208–210. doi: 10.1016/j.elecom.2013.06.016
|
| [31] |
Adler SB (2002) Reference Electrode Placement in Thin Solid Electrolytes. J Electrochem Soc 149: E166–E172. doi: 10.1149/1.1467368
|
| [32] |
Klink S, Höche D, La Mantia F, et al. (2013) FEM modelling of a coaxial three-electrode test cell for electrochemical impedance spectroscopy in lithium ion batteries. J Power Sources 240: 273–280. doi: 10.1016/j.jpowsour.2013.03.186
|
| [33] |
Momma T, Yokoshima T, Nara H, et al. (2014) Distinction of impedance responses of Li-ion batteries for individual electrodes using symmetric cells. Electrochimica Acta 131: 195–201. doi: 10.1016/j.electacta.2014.01.091
|
| [34] | Chen CH, Liu J, Amine K (2001) Symmetric cell approach and impedance spectroscopy of high power lithium-ion batteries. J Power Sources 96: 321–328. |
| [35] |
Levi MD, Dargel V, Shilina Y, et al. (2014) Impedance Spectra of Energy-Storage Electrodes Obtained with Commercial Three-Electrode Cells: Some Sources of Measurement Artefacts. Electrochimica Acta 149: 126–135. doi: 10.1016/j.electacta.2014.10.083
|
| [36] | Amatucci G (1996) Cobalt dissolution in LiCoO2-based non-aqueous rechargeable batteries. Solid State Ionics 83: 167–173. |
| [37] | Levi MD, Salitra G, Markovsky B, et al. (1999) Solid‐State Electrochemical Kinetics of Li‐Ion Intercalation into Li1 − x CoO2: Simultaneous Application of Electroanalytical Techniques SSCV, PITT, and EIS. J Electrochem Soc 146: 1279–1289. |
| [38] |
Momma T, Matsunaga M, Mukoyama D, et al. (2012) Ac impedance analysis of lithium ion battery under temperature control. J Power Sources 216: 304–307. doi: 10.1016/j.jpowsour.2012.05.095
|
| [39] | Nara H, Mukoyama D, Yokoshima T, et al. (2016) Impedance Analysis with Transmission Line Model for Reaction Distribution in a Pouch Type Lithium-Ion Battery by Using Micro Reference Electrode. J Electrochem Soc 163: A434–A441. |
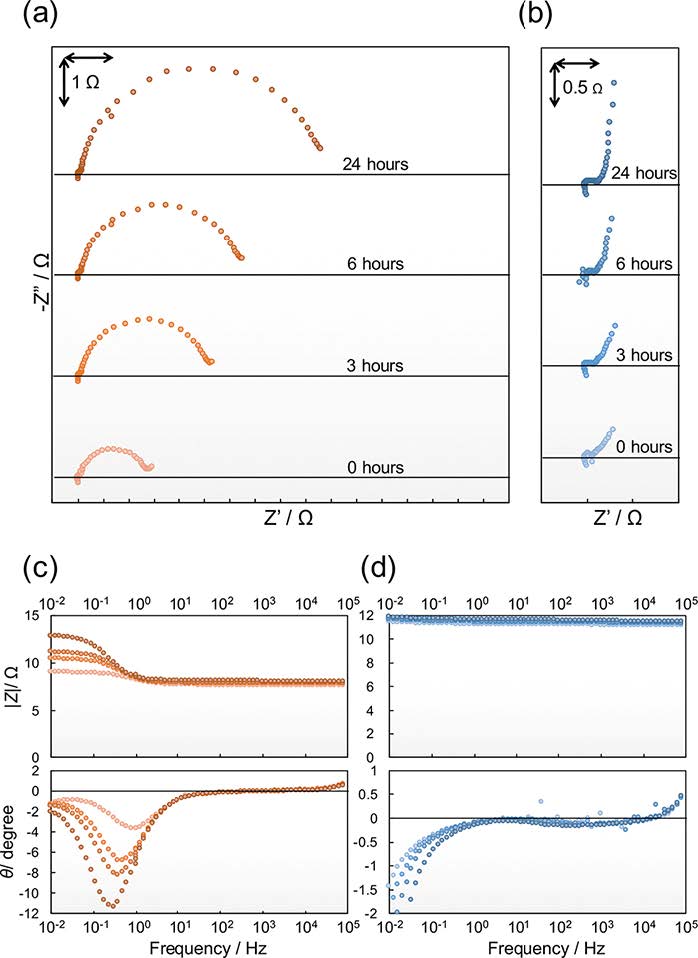
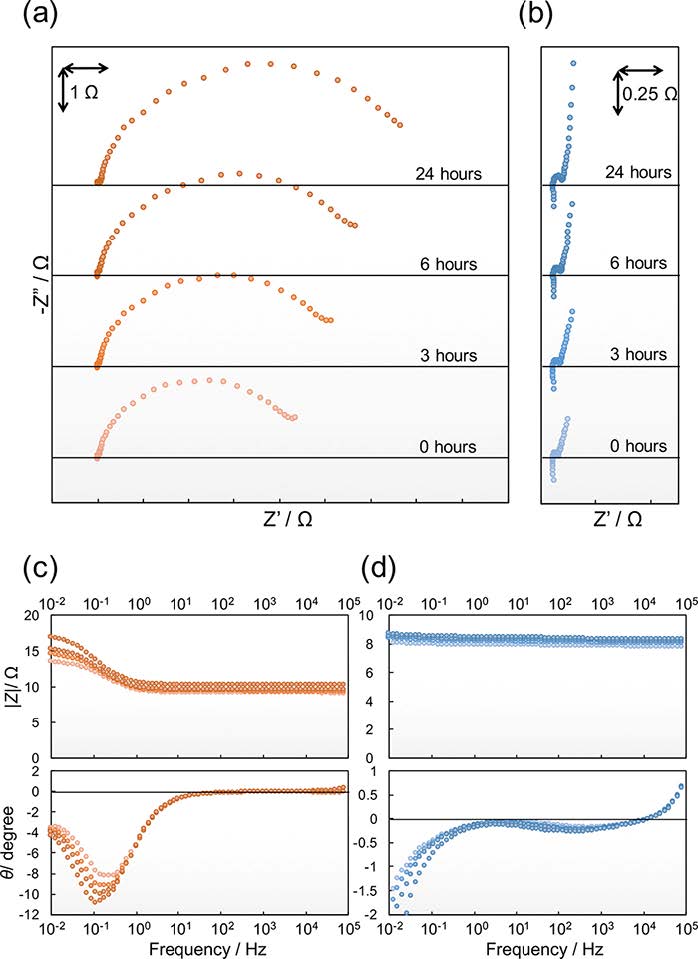
Figures(6)

Hiroki Nara, Keisuke Morita, Tokihiko Yokoshima, Daikichi Mukoyama, Toshiyuki Momma, Tetsuya Osaka. Electrochemical impedance spectroscopy analysis with a symmetric cell for LiCoO2 cathode degradation correlated with Co dissolution[J]. AIMS Materials Science, 2016, 3(2): 448-459. doi: 10.3934/matersci.2016.2.448







 DownLoad:
DownLoad: