Citation: Chiara De Santi, Sucharitha Gadi, Agnieszka Swiatecka-Urban, Catherine M. Greene. Identification of a novel functional miR-143-5p recognition element in the Cystic Fibrosis Transmembrane Conductance Regulator 3’UTR[J]. AIMS Genetics, 2018, 5(1): 53-62. doi: 10.3934/genet.2018.1.53

| [1] | Lee RC, Ambros V (1993) An extensive class of small RNAs in Caenorhabditis elegans. Science 294: 862–864. |
| [2] |
Chandra S, Vimal D, Sharma D, et al. (2017) Role of miRNAs in development and disease: Lessons learnt from small organisms. Life Sci 185: 8–14. doi: 10.1016/j.lfs.2017.07.017

|
| [3] |
Rupaimoole R, Calin GA, Lopez-Berestein G, et al. (2016) miRNA Deregulation in Cancer Cells and the Tumor Microenvironment. Cancer Discov 6: 235–246. doi: 10.1158/2159-8290.CD-15-0893
|
| [4] | Szymczak I, Wieczfinska J, Pawliczak R (2016) Molecular Background of miRNA Role in Asthma and COPD: An Updated Insight. Biomed Res Int 2016: 7802521. |
| [5] |
Chen JQ, Papp G, Szodoray P (2016) The role of microRNAs in the pathogenesis of autoimmune diseases. Autoimmun Rev 15: 1171–1180. doi: 10.1016/j.autrev.2016.09.003
|
| [6] |
Kiriakidou M, Nelson PT, Kouranov A, et al. (2004) A combined computational-experimental approach predicts human microRNA targets. Genes Dev 18: 1165–1178. doi: 10.1101/gad.1184704
|
| [7] |
Lai EC, Tam B, Rubin GM (2005) Pervasive regulation of Drosophila Notch target genes by GY-box-, Brd-box-, and K-box-class microRNAs. Genes Dev 19: 1067–1080. doi: 10.1101/gad.1291905
|
| [8] |
Vodicka P, Pardini B, Vymetalkova V, et al. (2016) Polymorphisms in Non-coding RNA Genes and Their Targets Sites as Risk Factors of Sporadic Colorectal Cancer. Adv Exp Med Biol 937: 123–149. doi: 10.1007/978-3-319-42059-2_7
|
| [9] |
Nossent AY, Hansen JL, Doggen C, et al. (2011) SNPs in microRNA binding sites in 3'-UTRs of RAAS genes influence arterial blood pressure and risk of myocardial infarction. Am J Hypertens 24: 999–1006. doi: 10.1038/ajh.2011.92
|
| [10] | Wang X, Jiang H, Wu W, et al. (2017) An Integrating Approach for Genome-Wide Screening of MicroRNA Polymorphisms Mediated Drug Response Alterations. Int J Genomics 2017: 1674827. |
| [11] |
Thomson DW, Bracken CP, Goodall GJ (2011) Experimental strategies for microRNA target identification. Nucleic Acids Res 39: 6845–6853. doi: 10.1093/nar/gkr330
|
| [12] |
Gillen AE, Gosalia N, Leir SH, et al. (2011) MicroRNA regulation of expression of the cystic fibrosis transmembrane conductance regulator gene. Biochem J 438: 25–32. doi: 10.1042/BJ20110672
|
| [13] |
Megiorni F, Cialfi S, Dominici C, et al. (2011) Synergistic post-transcriptional regulation of the Cystic Fibrosis Transmembrane conductance Regulator (CFTR) by miR-101 and miR-494 specific binding. PLoS One 6: e26601. doi: 10.1371/journal.pone.0026601
|
| [14] |
Hassan F, Nuovo GJ, Crawford M, et al. (2012) MiR-101 and miR-144 regulate the expression of the CFTR chloride channel in the lung. PLoS One 7: e50837. doi: 10.1371/journal.pone.0050837
|
| [15] |
Oglesby IK, Chotirmall SH, McElvaney NG, et al. (2013) Regulation of cystic fibrosis transmembrane conductance regulator by microRNA-145, -223, and -494 is altered in ΔF508 cystic fibrosis airway epithelium. J Immunol 190: 3354–3362. doi: 10.4049/jimmunol.1202960
|
| [16] |
Amato F, Seia M, Giordano S, et al. (2013) Gene mutation in microRNA target sites of CFTR gene: a novel pathogenetic mechanism in cystic fibrosis? PLoS One 8: e60448. doi: 10.1371/journal.pone.0060448
|
| [17] |
Ramachandran S, Karp PH, Osterhaus SR, et al. (2013) Post-transcriptional regulation of cystic fibrosis transmembrane conductance regulator expression and function by microRNAs. Am J Respir Cell Mol Biol 49: 544–551. doi: 10.1165/rcmb.2012-0430OC
|
| [18] |
Viart V, Bergougnoux A, Bonini J, et al. (2015) Transcription factors and miRNAs that regulate fetal to adult CFTR expression change are new targets for cystic fibrosis. Eur Respir J 45: 116–128. doi: 10.1183/09031936.00113214
|
| [19] | World Health Organization. The molecular genetic epidemiology of cystic fibrosis: report of a joint meeting of WHO/ECFTN/ICF(M)A/ECFS; June 19, 2002; Genoa, Italy. |
| [20] | The Clinical and Functional TRanslation of CFTR (CFTR2), 2017. Available from: http://cftr2.org. |
| [21] | McKiernan PJ, Greene CM (2015) MicroRNA Dysregulation in Cystic Fibrosis. Mediators Inflamm 2015: 529642. |
| [22] |
Oglesby IK, Bray IM, Chotirmall SH, et al. (2010) miR-126 is downregulated in cystic fibrosis airway epithelial cells and regulates TOM1 expression. J Immunol 184: 1702–1709. doi: 10.4049/jimmunol.0902669
|
| [23] |
Oglesby IK, Vencken SF, Agrawal R, et al. (2015) miR-17 overexpression in cystic fibrosis airway epithelial cells decreases interleukin-8 production. Eur Respir J 46: 1350–1360. doi: 10.1183/09031936.00163414
|
| [24] |
Mirković B, Murray MA, Lavelle GM, et al. (2015) The Role of Short-Chain Fatty Acids, Produced by Anaerobic Bacteria, in the Cystic Fibrosis Airway. Am J Respir Crit Care Med 192: 1314–1324. doi: 10.1164/rccm.201505-0943OC
|
| [25] |
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402–408. doi: 10.1006/meth.2001.1262
|
| [26] | Agarwal V, Bell GW, Nam JW, et al. (2015) Predicting effective microRNA target sites in mammalian mRNAs. Elife 4. |
| [27] |
Betel D, Koppal A, Agius P, et al. (2010) Comprehensive modeling of microRNA targets predicts functional non-conserved and non-canonical sites. Genome Biol 11: R90. doi: 10.1186/gb-2010-11-8-r90
|
| [28] |
Miranda KC, Huynh T, Tay Y, et al. (2006) A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 126: 1203–1217. doi: 10.1016/j.cell.2006.07.031
|
| [29] |
Clément T, Salone V, Rederstorff M (2015) Dual luciferase gene reporter assays to study miRNA function. Methods Mol Biol 1296: 187–198. doi: 10.1007/978-1-4939-2547-6_17
|
| [30] |
Karimi L, Mansoori B, Shanebandi D, et al. (2017) Function of microRNA-143 in different signal pathways in cancer: New insights into cancer therapy. Biomed Pharmacother 91: 121–131. doi: 10.1016/j.biopha.2017.04.060
|
| [31] |
He M, Zhan M, Chen W, et al. (2017) MiR-143-5p Deficiency Triggers EMT and Metastasis by Targeting HIF-1α in Gallbladder Cancer. Cell Physiol Biochem 42: 2078–2092. doi: 10.1159/000479903
|
| [32] |
Wu XL, Cheng B, Li PY, et al. (2013) MicroRNA-143 suppresses gastric cancer cell growth and induces apoptosis by targeting COX-2. World J Gastroenterol 19: 7758–7765. doi: 10.3748/wjg.v19.i43.7758
|
| [33] | FDA expands approved use of Kalydeco to treat additional mutations of cystic fibrosis, FDA News Release, 2017. Available from: https://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm559212.htm |
| [34] |
Fajac I, De Boeck K (2017) New horizons for cystic fibrosis treatment. Pharmacol Ther 170: 205–211. doi: 10.1016/j.pharmthera.2016.11.009
|
| [35] |
Clancy JP, Rowe SM, Accurso FJ, et al. (2012) Results of a phase IIa study of VX-809, an investigational CFTR corrector compound, in subjects with cystic fibrosis homozygous for the F508del-CFTR mutation. Thorax 67: 12–18. doi: 10.1136/thoraxjnl-2011-200393
|
| [36] |
Boyle MP, Bell SC, Konstan MW, et al. (2014) A CFTR corrector (lumacaftor) and a CFTR potentiator (ivacaftor) for treatment of patients with cystic fibrosis who have a phe508del CFTR mutation: a phase 2 randomised controlled trial. Lancet Respir Med 2: 527–538. doi: 10.1016/S2213-2600(14)70132-8
|
| [37] |
Wainwright CE, Elborn JS, Ramsey BW, et al. (2015) Lumacaftor-Ivacaftor in Patients with Cystic Fibrosis Homozygous for Phe508del CFTR. N Engl J Med 373: 220–231. doi: 10.1056/NEJMoa1409547
|
| [38] |
Zarrilli F, Amato F, Morgillo CM, et al. (2017) Peptide Nucleic Acids as miRNA Target Protectors for the Treatment of Cystic Fibrosis. Molecules 22: 1144. doi: 10.3390/molecules22071144
|
| [39] | Cantin AM (2016) Cystic Fibrosis Transmembrane Conductance Regulator. Implications in Cystic Fibrosis and Chronic Obstructive Pulmonary Disease. Ann Am Thorac Soc 13 Suppl 2: S150–S155. |
| [40] | Solomon GM, Raju SV, Dransfield MT, et al. (2016) Therapeutic Approaches to Acquired Cystic Fibrosis Transmembrane Conductance Regulator Dysfunction in Chronic Bronchitis. Ann Am Thorac Soc 13 Suppl 2: S169–S176. |
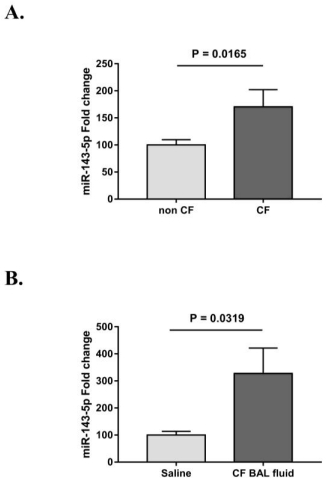
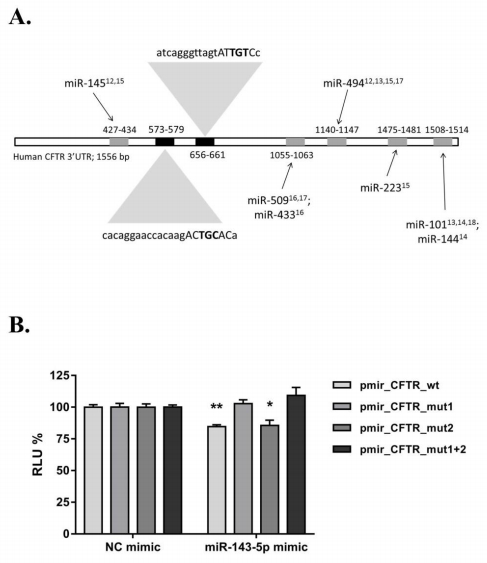
Figures(2)

Chiara De Santi, Sucharitha Gadi, Agnieszka Swiatecka-Urban, Catherine M. Greene. Identification of a novel functional miR-143-5p recognition element in the Cystic Fibrosis Transmembrane Conductance Regulator 3’UTR[J]. AIMS Genetics, 2018, 5(1): 53-62. doi: 10.3934/genet.2018.1.53







 DownLoad:
DownLoad: