Citation: Noel Pabalan, Neetu Singh, Eloisa Singian, Caio Parente Barbosa, Bianca Bianco, Hamdi Jarjanazi. Associations of CYP1A1 gene polymorphisms and risk of breast cancer in Indian women: a meta-analysis[J]. AIMS Genetics, 2015, 2(4): 250-262. doi: 10.3934/genet.2015.4.250

| [1] |
Dikshit R, Gupta PC, Ramasundarahettige C, et al. (2012) Cancer mortality in India: a nationally representative survey. Lancet 379: 1807-1816. doi: 10.1016/S0140-6736(12)60358-4

|
| [2] |
Vargo-Gogola T, Rosen JM (2007) Modelling breast cancer: one size does not fit all. Nat Rev Cancer 7: 659-672. doi: 10.1038/nrc2193
|
| [3] |
Nickels S, Truong T, Hein R, et al. (2013) Evidence of gene-environment interactions between common breast cancer susceptibility loci and established environmental risk factors. PLoS Genet 9: e1003284. doi: 10.1371/journal.pgen.1003284
|
| [4] |
Chen C, Huang Y, Li Y, et al. (2007) Cytochrome P450 1A1 (CYP1A1) T3801C and A2455G polymorphisms in breast cancer risk: a meta-analysis. J Hum Genet 52: 423-435. doi: 10.1007/s10038-007-0131-8
|
| [5] |
Sergentanis TN, Economopoulos KP (2010) Four polymorphisms in cytochrome P450 1A1 (CYP1A1) gene and breast cancer risk: a meta-analysis. Breast Cancer Res Treat 122: 459-469. doi: 10.1007/s10549-009-0694-5
|
| [6] |
Yao L, Yu X, Yu L (2010) Lack of significant association between CYP1A1 T3801C polymorphism and breast cancer risk: a meta-analysis involving 25,087 subjects. Breast Cancer Res Treat 122: 503-507. doi: 10.1007/s10549-009-0717-2
|
| [7] |
Syamala VS, Syamala V, Sheeja VR, et al. (2010) Possible risk modification by polymorphisms of estrogen metabolizing genes in familial breast cancer susceptibility in an Indian population. Cancer Invest 28: 304-311. doi: 10.3109/07357900902744494
|
| [8] |
Surekha D, Sailaja K, Rao DN, et al. (2009) Association of CYP1A1*2 polymorphisms with breast cancer risk: a case control study. Indian J Med Sci 63: 13-20. doi: 10.4103/0019-5359.49077
|
| [9] |
Singh V, Rastogi N, Sinha A, et al. (2007) A study on the association of cytochrome-P450 1A1 polymorphism and breast cancer risk in north Indian women. Breast Cancer Res Treat 101: 73-81. doi: 10.1007/s10549-006-9264-2
|
| [10] |
Singh N, Mitra AK, Garg VK, et al. (2007) Association of CYP1A1 polymorphisms with breast cancer in North Indian women. Oncol Res 16: 587-597. doi: 10.3727/000000007783629972
|
| [11] |
Naushad SM, Reddy CA, Rupasree Y, et al. (2011) Cross-talk between one-carbon metabolism and xenobiotic metabolism: implications on oxidative DNA damage and susceptibility to breast cancer. Cell Biochem Biophys 61: 715-723. doi: 10.1007/s12013-011-9245-x
|
| [12] |
Chacko P, Joseph T, Mathew BS, et al. (2005) Role of xenobiotic metabolizing gene polymorphisms in breast cancer susceptibility and treatment outcome. Mutat Res 581: 153-163. doi: 10.1016/j.mrgentox.2004.11.018
|
| [13] | Kiruthiga PV, Kannan MR, Saraswathi C, et al. (2011) CYP1A1 gene polymorphisms: lack of association with breast cancer susceptibility in the southern region (Madurai) of India. Asian Pac J Cancer Prev 12: 2133-2138. |
| [14] |
Masson LF, Sharp L, Cotton SC, et al. (2005) Cytochrome P-450 1A1 gene polymorphisms and risk of breast cancer: a HuGE review. Am J Epidemiol 161: 901-915. doi: 10.1093/aje/kwi121
|
| [15] |
Kawajiri K, Nakachi K, Imai K, et al. (1990) Identification of genetically high risk individuals to lung cancer by DNA polymorphisms of the cytochrome P450IA1 gene. FEBS Lett 263: 131-133. doi: 10.1016/0014-5793(90)80721-T
|
| [16] | Hayashi SI, Watanabe J, Nakachi K, et al. (1991) PCR detection of an A/G polymorphism within exon 7 of the CYP1A1 gene. Nucleic Acids Res 19: 4797. |
| [17] | Cascorbi I, Brockmoller J, Roots I (1996) A C4887A polymorphism in exon 7 of human CYP1A1: population frequency, mutation linkages, and impact on lung cancer susceptibility. Cancer Res 56: 4965-4969. |
| [18] | Li Y, Millikan RC, Bell DA, et al. (2005) Polychlorinated biphenyls, cytochrome P450 1A1 (CYP1A1) polymorphisms, and breast cancer risk among African American women and white women in North Carolina: a population-based case-control study. Breast Cancer Res 7: R12-18. |
| [19] |
Crofts F, Taioli E, Trachman J, et al. (1994) Functional significance of different human CYP1A1 genotypes. Carcinogenesis 15: 2961-2963. doi: 10.1093/carcin/15.12.2961
|
| [20] |
Kiyohara C, Hirohata T, Inutsuka S (1996) The relationship between aryl hydrocarbon hydroxylase and polymorphisms of the CYP1A1 gene. Jpn J Cancer Res 87: 18-24. doi: 10.1111/j.1349-7006.1996.tb00194.x
|
| [21] |
Li Y, Millikan RC, Bell DA, et al. (2004) Cigarette smoking, cytochrome P4501A1 polymorphisms, and breast cancer among African-American and white women. Breast Cancer Res 6: R460-473. doi: 10.1186/bcr814
|
| [22] |
Moher D, Liberati A, Tetzlaff J, et al. (2009) Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. Ann Intern Med 151: 264-269, W264. doi: 10.7326/0003-4819-151-4-200908180-00135
|
| [23] | Wells S PJ, Welch V. The newcastle-ottawa scale (NOS) for assessing the quality of nonrandomised studies in meta-analyses. Ottawa Health Research Institute, 2011. Available from: www.ohri.ca/programs/clinical_epidemiology/oxford/asp. |
| [24] | Mantel N, Haenszel W (1959) Statistical aspects of the analysis of data from retrospective studies of disease. J Natl Cancer Inst 22: 719-748. |
| [25] |
DerSimonian R, Laird N (1986) Meta-analysis in clinical trials. Control Clin Trials 7: 177-188. doi: 10.1016/0197-2456(86)90046-2
|
| [26] |
Lau J, Ioannidis JP, Schmid CH (1997) Quantitative synthesis in systematic reviews. Ann Intern Med 127: 820-826. doi: 10.7326/0003-4819-127-9-199711010-00008
|
| [27] |
Higgins JP, Thompson SG (2002) Quantifying heterogeneity in a meta-analysis. Stat Med 21: 1539-1558. doi: 10.1002/sim.1186
|
| [28] |
Higgins JP, Thompson SG, Deeks JJ, et al. (2003) Measuring inconsistency in meta-analyses. Bmj 327: 557-560. doi: 10.1136/bmj.327.7414.557
|
| [29] | Gautham M, Shyamprasad KM, Singh R, et al. (2014) Informal rural healthcare providers in North and South India. Health Policy Plan 29 Suppl 1: i20-29. |
| [30] |
Ravindran RD, Vashist P, Gupta SK, et al. (2011) Prevalence and risk factors for vitamin C deficiency in north and south India: a two centre population based study in people aged 60 years and over. PLoS One 6: e28588. doi: 10.1371/journal.pone.0028588
|
| [31] |
Ioannidis JP, Trikalinos TA (2007) The appropriateness of asymmetry tests for publication bias in meta-analyses: a large survey. Cmaj 176: 1091-1096. doi: 10.1503/cmaj.060410
|
| [32] | Gadgil MaG, R. (1992) The fissure land: An ecological history of India; Press OU, editor. New Delhi: Oxford University Press. |
| [33] |
He XF, Wei W, Liu ZZ, et al. (2014) Association between the CYP1A1 T3801C polymorphism and risk of cancer: evidence from 268 case-control studies. Gene 534: 324-344. doi: 10.1016/j.gene.2013.10.025
|
| [34] |
Wacholder S, Chanock S, Garcia-Closas M, et al. (2004) Assessing the probability that a positive report is false: an approach for molecular epidemiology studies. J Natl Cancer Inst 96: 434-442. doi: 10.1093/jnci/djh075
|
| [35] |
Thakkinstian A, McElduff P, D'Este C, et al. (2005) A method for meta-analysis of molecular association studies. Stat Med 24: 1291-1306. doi: 10.1002/sim.2010
|
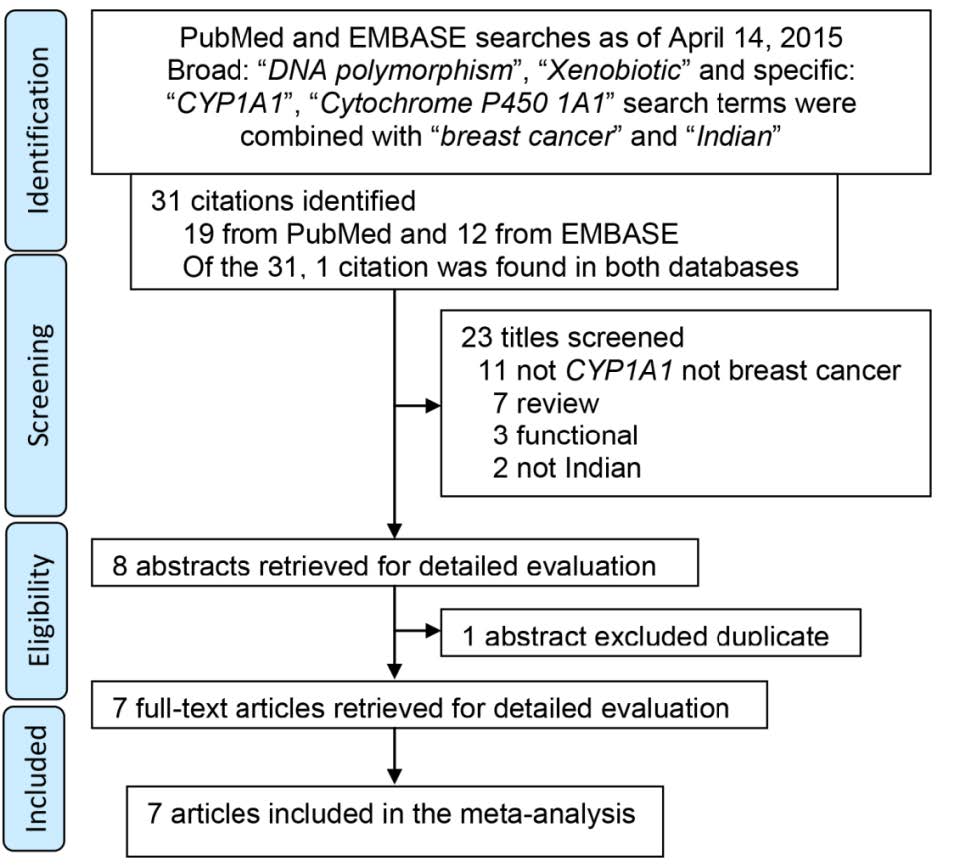
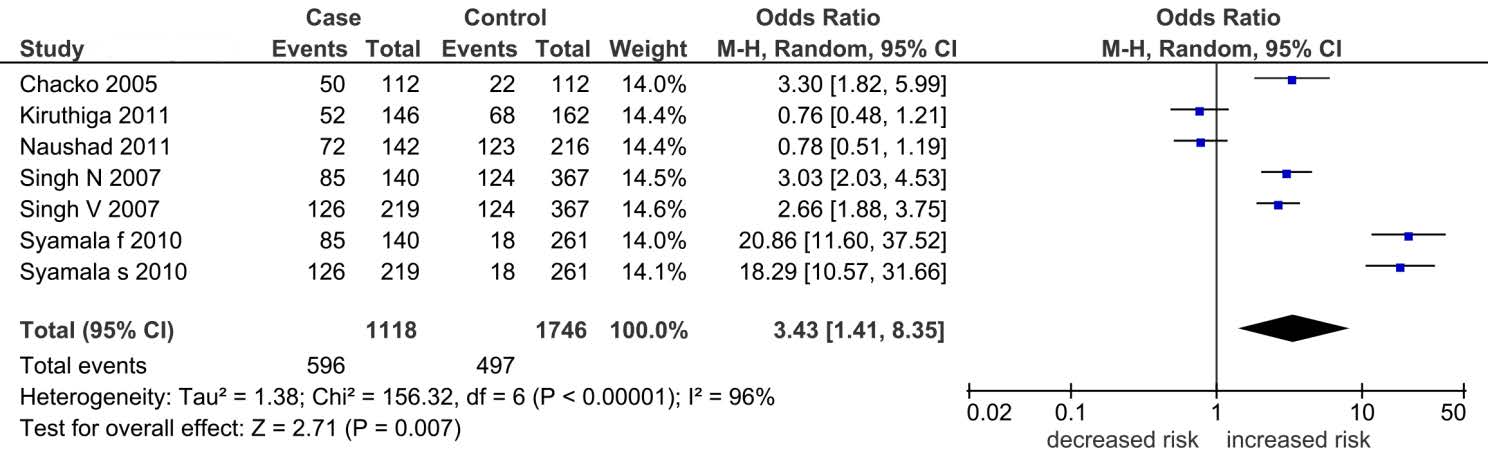
Figures(2) / Tables(7)

Noel Pabalan, Neetu Singh, Eloisa Singian, Caio Parente Barbosa, Bianca Bianco, Hamdi Jarjanazi. Associations of CYP1A1 gene polymorphisms and risk of breast cancer in Indian women: a meta-analysis[J]. AIMS Genetics, 2015, 2(4): 250-262. doi: 10.3934/genet.2015.4.250







 DownLoad:
DownLoad: