Citation: Natalie J. Nokoff, Sharon Travers, Naomi Meeks. Two cases of androgen insensitivity due to somatic mosaicism[J]. AIMS Genetics, 2015, 2(2): 104-109. doi: 10.3934/genet.2015.2.104

| [1] | Gottlieb B, Beitel LK, Trifiro MA (1999 Mar 24 Updated 2014 Jul 10.) Androgen Insensitivity Syndrome. GeneReviews® Internet. Seattle (WA): University of Washington, Seattle; 1993-2014 Available from: http://wwwncbinlmnihgov/books/NBK1429/. |
| [2] |
Galani A, Kitsiou-Tzeli S, Sofokleous C, et al. (2008) Androgen insensitivity syndrome: clinical features and molecular defects. Hormones 7: 217-229. doi: 10.14310/horm.2002.1201

|
| [3] |
Boehmer ALM, Brüggenwirth H, van Assendelft C, et al. (2001) Genotype Versus Phenotype in Families with Androgen Insensitivity Syndrome. J Clin Endocrinol Metab 86: 4151-4160. doi: 10.1210/jcem.86.9.7825
|
| [4] |
Hiort O, Sinnecker GH, Holterhus PM, et al. (1998) Inherited and de novo androgen receptor gene mutations: investigation of single-case families. J Pediatr 132: 939-943. doi: 10.1016/S0022-3476(98)70387-7
|
| [5] | Holterhus PM, Bruggenwirth HT, Hiort O, et al. (1997) Mosaicism due to a somatic mutation of the androgen receptor gene determines phenotype in androgen insensitivity syndrome. J Clin Endocrinol Metab 82: 3584-3589. |
| [6] |
Holterhus P-M, Wiebel J, Sinnecker GHG, et al. (1999) Clinical and Molecular Spectrum of Somatic Mosaicism in Androgen Insensitivity Syndrome. Pediatr Res 46: 684-684. doi: 10.1203/00006450-199912000-00009
|
| [7] | Boehmer AL, Brinkmann AO, Niermeijer MF, et al. (1997) Germ-line and somatic mosaicism in the androgen insensitivity syndrome: implications for genetic counseling. Am J Hum Genet 60: 1003-1006. |
| [8] |
Köhler B, Lumbroso S, Leger J, et al. (2005) Androgen insensitivity syndrome: somatic mosaicism of the androgen receptor in seven families and consequences for sex assignment and genetic counseling. J Clin Endocrinol Metab 90: 106-111. doi: 10.1210/jc.2004-0462
|
| [9] |
Hughes IA, Werner R, Bunch T, et al. (2012) Androgen insensitivity syndrome. Semin Reprod Med 30: 432-442. doi: 10.1055/s-0032-1324728
|
| [10] |
Gottlieb B, Beitel LK, Nadarajah A, et al. (2012) The androgen receptor gene mutations database: 2012 update. Human Mutation 33: 887-894. doi: 10.1002/humu.22046
|
| [11] | Sperling M (2014) Pediatric endocrinology fourth edition. Philadelphia, PA: Saunders/Elsevier. |
| [12] |
Gottlieb B, Beitel LK, Trifiro MA (2001) Somatic mosaicism and variable expressivity. Trends Genet 17: 79-82. doi: 10.1016/S0168-9525(00)02178-8
|
| [13] |
Hook EB, Warburton D (2014) Turner syndrome revisited: review of new data supports the hypothesis that all viable 45,X cases are cryptic mosaics with a rescue cell line, implying an origin by mitotic loss. Hum Genet 133: 417-424. doi: 10.1007/s00439-014-1420-x
|
| [14] |
Zoppi S, Wilson CM, Harbison MD, et al. (1993) Complete testicular feminization caused by an amino-terminal truncation of the androgen receptor with downstream initiation. J Clin Invest 91: 1105-1112. doi: 10.1172/JCI116269
|
| [15] |
Trifiro M, Prior RL, Sabbaghian N, et al. (1991) Amber mutation creates a diagnostic MaeI site in the androgen receptor gene of a family with complete androgen insensitivity. Am J Med Genet 40: 493-499. doi: 10.1002/ajmg.1320400425
|
| [16] | Hiort O, Wodtke A, Struve D, et al. (1994) Detection of point mutations in the androgen receptor gene using non-isotopic single strand conformation polymorphism analysis. German Collaborative Intersex Study Group. Hum Mol Genet 3: 1163-1166. |
| [17] |
Batch JA, Williams DM, Davies HR, et al. (1992) Androgen receptor gene mutations identified by SSCP in fourteen subjects with androgen insensitivity syndrome. Hum Mol Genet 1: 497-503. doi: 10.1093/hmg/1.7.497
|
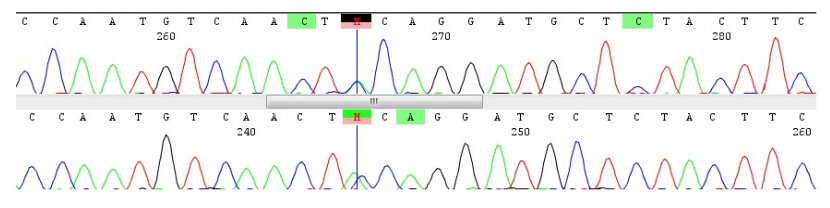
Figures(1)

Natalie J. Nokoff, Sharon Travers, Naomi Meeks. Two cases of androgen insensitivity due to somatic mosaicism[J]. AIMS Genetics, 2015, 2(2): 104-109. doi: 10.3934/genet.2015.2.104







 DownLoad:
DownLoad: