Citation: Espen Olsen, Maria Hansen, Heidi S. Nygård. Hydrolysis of molten CaCl2-CaF2 with additions of CaO[J]. AIMS Energy, 2017, 5(6): 873-886. doi: 10.3934/energy.2017.6.873

| [1] |
Chu S (2009) Carbon capture and sequestration. Science 325: 1599. doi: 10.1126/science.1181637

|
| [2] |
Macdowell N, Florin N, Buchard A, et al. (2010) An overview of CO2 capture technologies. Energ Environ Sci 3: 1645–1669. doi: 10.1039/c004106h
|
| [3] |
Dean CC, Blamey J, Florin NH, et al. (2011) The calcium looping cycle for CO2 capture from power generation, cement manufacture and hydrogen production. Chem Eng Res Des 89: 836–855. doi: 10.1016/j.cherd.2010.10.013
|
| [4] | Junk M, Reitz M, Strohle J, et al. (2016) Technical and economical assessment of the indirectly heated carbonate looping process. J Energ Resour Technol 138. |
| [5] |
Abanades JC, Alvarez D (2003) Conversion limits in the reaction of CO2 with lime. Energ Fuel 17: 308–315. doi: 10.1021/ef020152a
|
| [6] |
Donat F, Florin NH, Anthony EJ, et al. (2012) Influence of high-temperature steam on the reactivity of CaO sorbent for CO2 capture. Environ Sci Technol 46: 1262–1269. doi: 10.1021/es202679w
|
| [7] |
Aljeboori MJ, Nguyen M, Dean C, et al. (2013) Improvement of limestone-based CO2 sorbents for Ca looping by HBr and other mineral acids. Ind Eng Chem Res 52: 1426–1433. doi: 10.1021/ie302198g
|
| [8] |
Liu W, Yin J, Qin C, et al. (2012) Synthesis of CaO-based sorbents for CO2 capture by a spray-drying technique. Environ Sci Technol 46: 11267–11272. doi: 10.1021/es301783b
|
| [9] |
Stendardo S, Andersen LK, Herce C (2013) Self-activation and effect of regeneration conditions in CO2-carbonate looping with CaO-Ca12Al14O33 sorbent. Chem Eng J 220: 383–394. doi: 10.1016/j.cej.2013.01.045
|
| [10] |
Olsen E, Tomkute V (2013) Carbon capture in molten salts. Energy Sci Eng 1: 144–150. doi: 10.1002/ese3.24
|
| [11] | Tomkute V, Solheim A, Olsen E (2013) Investigation of high-temperature CO2 capture by CaO in CaCl2 molten salt. Energ Fuel 27: 5373–5379. |
| [12] |
Tomkute V, Solheim A, Olsen E (2014) CO2 capture by CaO in molten CaF2-CaCl2: optimization of the process and cyclability of CO2 capture. Energ Fuel 28: 5345–5353. doi: 10.1021/ef5010896
|
| [13] |
Chen GZ, Fray DJ, Farthing TW (2000) Direct electrochemical reduction of titanium dioxide to titanium in molten calcium chloride. Nature 407: 361–364. doi: 10.1038/35030069
|
| [14] |
Chen GZ, Fray DJ (2006) A morphological study of the FFC chromium and titanium powders. Miner Process Extr Metall Imm Trans 115: 49–54. doi: 10.1179/174328506X91365
|
| [15] |
Vishnu DSM, Sanil N, Mohandas KS, et al. (2017) Factors influencing the direct electrochemical reduction of Nb2O5 pellets to Nb metal in molten chloride salts. Acta Metall Sin 30: 218–227. doi: 10.1007/s40195-016-0503-1
|
| [16] | Kipouros GJ, Sadoway DR (2001) A thermochemical analysis of the production of anhydrous MgCl2. J Light Metals 2: 111–117. |
| [17] | Roine A (2008) HSC Chemistry 6. 6.1 ed. Pori Finland. Outokumpu Research. |
| [18] |
Kondo H, Asaki Z, Kondo Y (1978) Hydrolysis of fused calcium chloride at high temperature. Metall Trans B 9: 477–483. doi: 10.1007/BF02654424
|
| [19] | Hanf NW. Nickel Segregation Research at Anglo American Research Laboratories. Part 2. Hydrogen Chloride Evolution in the Nickel Segregation system. In: Dor AA, editor. Proceedings of the Panel Discussion on Nickel Segregation, 1972 Feb.23, San Francisco CA. American Institute of Mining, Metallurgical, and Petroleum Engineers, 62–71. |
| [20] |
Wang S, Zhang F, Liu X, et al. (2008) CaO solubility and activity coefficient in molten salts CaCl2–x (x = 0, NaCl, KCl, SrCl2, BaCl2 and LiCl). Thermochim Acta 470: 105–107. doi: 10.1016/j.tca.2008.02.007
|
| [21] | Sonntag RE, Borgnakke C (2007) Introduction to Engineering Thermodynamics. Hoboken NJ : John Wiley & Sons, 489 pl. |
| [22] |
Freidina EB, Fray DJ (2000) Phase diagram of the system CaCl2–CaCO3. Thermochim Acta 351: 107–108. doi: 10.1016/S0040-6031(00)00420-2
|
| [23] | Shen VK, Siderius DW, Krekelberg WP, et al. (2010) ACerS-NIST Phase Equilibria Diagrams NIST Standard Reference Database, 3.1 ed., Westerville OH : The American Ceramic Society, 04908. |
| [24] |
Wenz DA, Johnson I, Wolson RD (1969) CaCl2-rich region of CaCl2-CaF2-CaO system. J Chem Eng Data 14: 250–251. doi: 10.1021/je60041a027
|
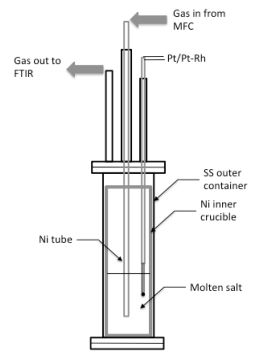
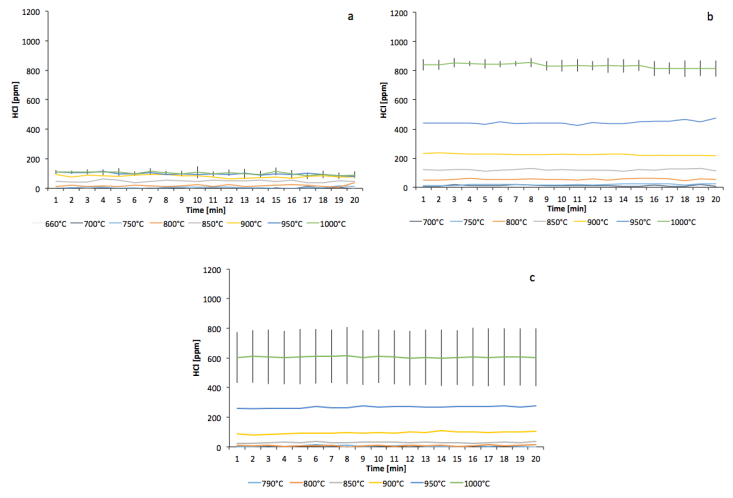
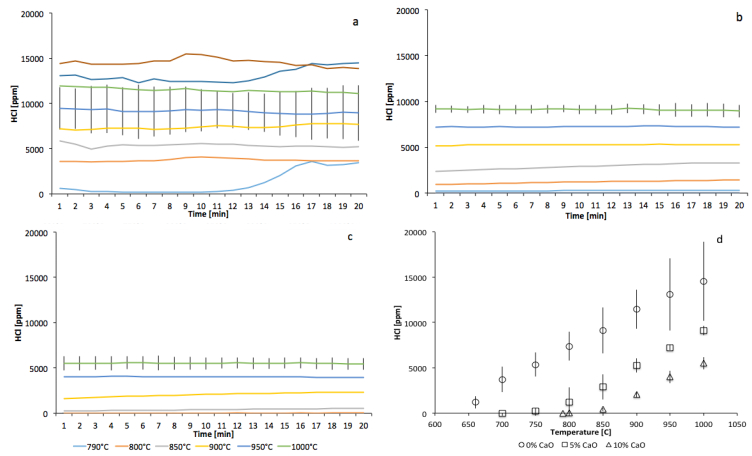

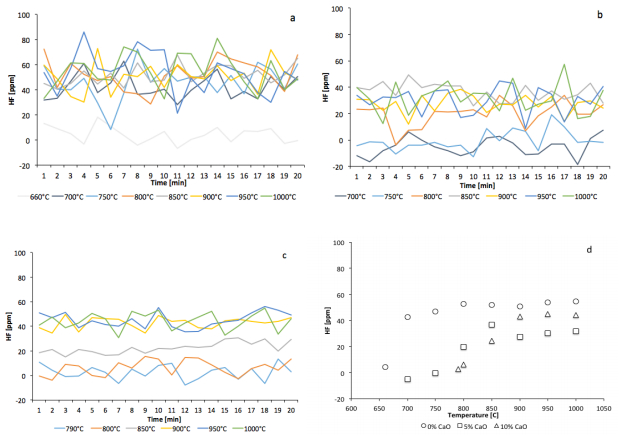
Figures(5) / Tables(3)

Espen Olsen, Maria Hansen, Heidi S. Nygård. Hydrolysis of molten CaCl2-CaF2 with additions of CaO[J]. AIMS Energy, 2017, 5(6): 873-886. doi: 10.3934/energy.2017.6.873







 DownLoad:
DownLoad: