Citation: G.Corradini Maria, LaviniaWang Yan, Le An, M.Waxman Sarah, Zelent Bogumil, Chib Rahul, Gryczynski Ignacy, D.Ludescher Richard. Identifying and selecting edible luminescent probes as sensors of food quality[J]. AIMS Biophysics, 2016, 3(2): 319-339. doi: 10.3934/biophy.2016.2.319

| [1] | Corradini MG, Ludescher RD (2015) Making sense of luminescence from GRAS optical probes. Curr Opin Food Sci 4: 25-31. |
| [2] | Wolfbeis OS (1985) The fluorescence of organic natural products. In Molecular luminescence spectroscopy: Methods and applications — Part 1 . Edited by Schulman SG: John Wiley and Sons, Toronto, Canada. |
| [3] | Christensen J, Norgaard L, Bro R, et al. (2006) Multivariate autofluorescence of intact food systems. Chem Rev 106: 1979-1994. |
| [4] | Karoui R, Mouazen AM, Dufour E, et al. (2006) Utilisation of front-face fluorescence spectroscopy for the determination of some selected chemical parameters in soft cheeses. Lait 86: 155-169. |
| [5] | Karoui R, Dufour E, De Baerdemaeker J (2007) Front face fluorescence spectroscopy coupled with chemometric tools for monitoring the oxidation of semi-hard cheeses throughout ripening. Food Chem 101: 1305-1314. |
| [6] | Karoui R, Blecker C (2011) Fluorescence spectroscopy measurement for quality assessment of food systems-a review. Food Bioprocess Tech 4: 364-386. |
| [7] | Botosoa EP, Chèné C, Karoui R (2013) Use of front face fluorescence for monitoring lipid oxidation during ageing of cakes. Food Chem 141: 1130-1139. |
| [8] | Airado-Rodriguez D, Hoy M, Skaret J, et al. (2014) From multispectral imaging of autofluorescence to chemical and sensory images of lipid oxidation in cod caviar paste. Talanta 122: 70-79. |
| [9] | Turro NJ, Ramamurthy V, Scaiano JC (2010) Modern Molecular Photochemistry of Organic Molecules . Sausalito, CA: University Science Books. |
| [10] | Chihara K, Takemura T, Yamaoka T, et al. (1979) Visual pigments—10. Spectroscopy and photophysical dynamics of retinol and retinyl ether. Photochem Photobiol 29: 1001-1008. |
| [11] | Shreve AP, Trautman JK, Owens TG, et al. (1991) Determination of the S2 lifetime of β-carotene. Chem Phys Lett 178: 89-96. |
| [12] | Rampon V, Lethuaut L, Mouhous-Riou N, et al. (2001) Interface characterization and aging of bovine serum albumin stabilized oil-in-water emulsions as revealed by front-surface fluorescence. J Agr Food Food Chem 49: 4046-4051. |
| [13] | Dufour E (2002) Examination of the molecular structure of food products using front-face fluorescence spectroscopy. Am Lab 34: 50. |
| [14] | Dufour E (2011) Recent advances in the analysis of dairy product quality using methods based on the interactions of light with matter. Int J Dairy Technol 64: 153-165. |
| [15] | Lakhal L, Acha V, Aussenac T (2014) Resolution of fluorophore mixtures in biological media using fluorescence spectroscopy and Monte Carlo simulation. Appl Spectrosc 68: 697-711. |
| [16] | Du H, Kim C, Corradini MG, et al. (2014) Micro-viscosity of liquid oil confined in colloidal fat crystal networks. Soft Matter 10: 8652-8658. |
| [17] | Bro R, Vidal M (2011) EEMizer: Automated modeling of fluorescence EEM data. Chemometr Intell Lab 106: 86-92. |
| [18] | Zhu C, Li R, Que L-z, et al. (2014) Determination of components of mixed color solutions by fluorescence spectroscopy combined with calibration algorithms. Spectrosc Spect Anal 34: 1742-1746. |
| [19] | Sadecka J, Tothova J (2007) Fluorescence spectroscopy and chemometrics in the food classification - a review. Czech J Food Sci 25: 159-173. |
| [20] | Skov T, Honore AH, Jensen HM, et al. (2014) Chemometrics in foodomics: Handling data structures from multiple analytical platforms. Trac-Trend Anal Chem 60: 71-79. |
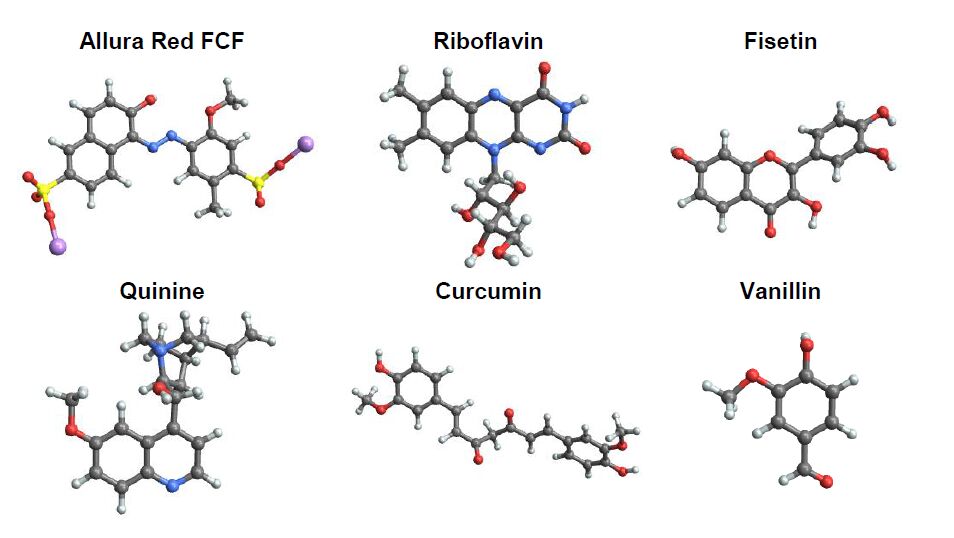
| [21] | Kashi A, Waxman SM, Komaiko JS, et al. (2015) Potential use of food synthetic colors as intrinsic luminescent probes of the physical state of foods. In Chemical Sensory Informatics of Food: Measurement, Analysis, Integration . Edited by Guthrie B, Buettner A, Beauchamp J, Lavine BK: ACS Publications - Oxford University Press; 253-267. |
| [22] | Nazar MF, Murtaza S (2014) Physicochemical investigation and spectral properties of Sunset Yellow dye in cetyltrimethylammonium bromide micellar solution under different pH conditions. Color Technol 130: 191-199. |
| [23] | Food, Administration D (2015) Code of Federal Regulations - Food Additives. Tittle 21 Chapter I: Part 74. |
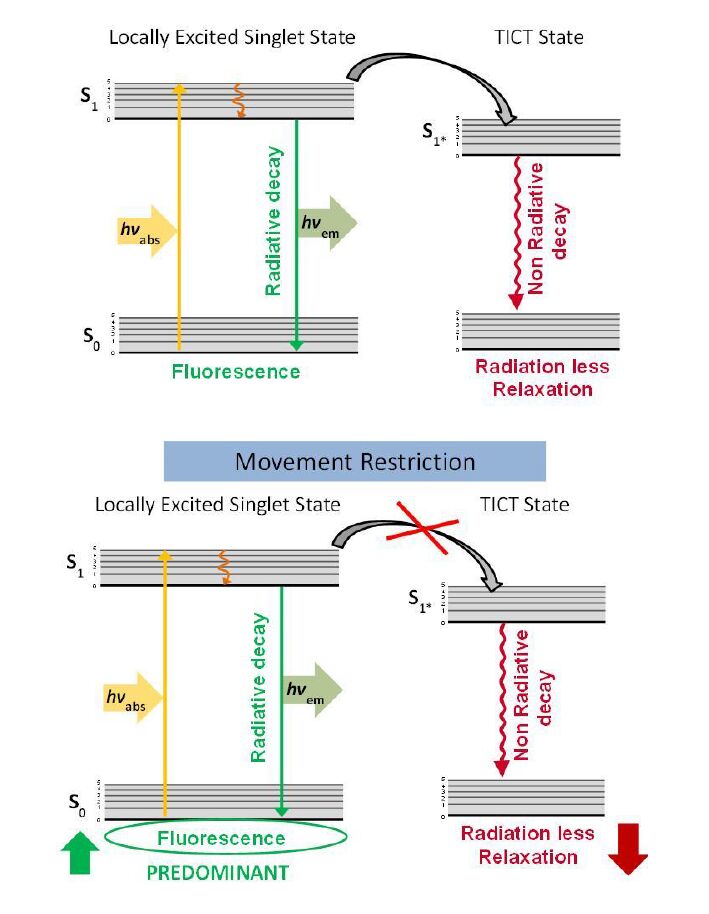
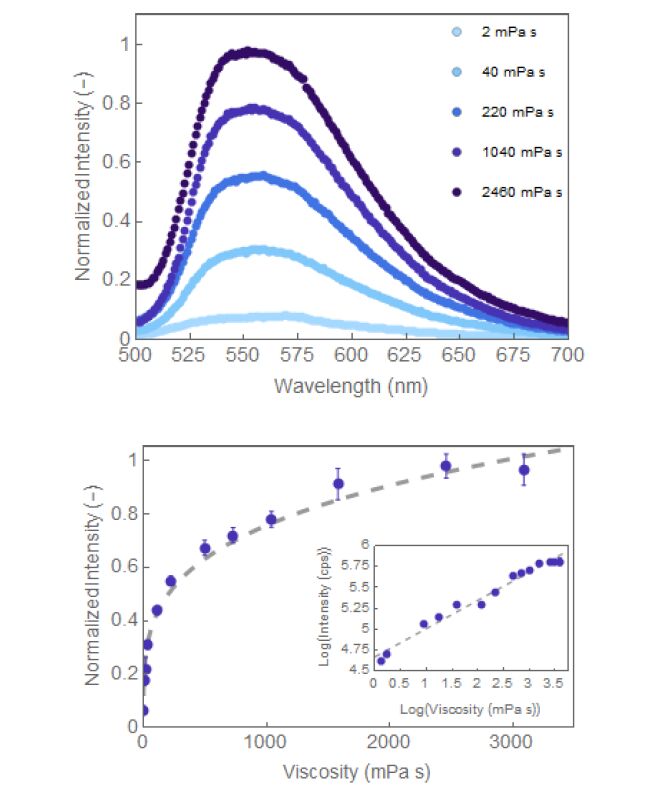
| [24] | Ludescher RD, Komaiko JS, Corradini MG (2012) Monoazo dyes as probes for bulk and local viscosity Potential applications in semi-solid foods. Agro Food Ind Hi-Tech 23: 11-13. |
| [25] | Law KY (1980) Fluorescence probe for microenvironments: anomalous viscosity dependence of the fluorescence quantum yield of p-N,N-dialkyl-amino-benzylidene-malononitrile in 1-alkanols. Chem Phys Lett 75: 545-549. |
| [26] | Chug S, Hanson JL, Ryoo S, et al. (2013) Potential application of natural pigments as probes for viscosity. In Abstracts of papers of the American Chemical Society : Washington, DC, USA. |
| [27] | Kuimova MK (2012) Molecular rotors image intracellularvViscosity. Chimia 66: 159-165. |
| [28] | Haidekker MA, Theodorakis EA (2010) Environment-sensitive behavior of fluorescent molecular rotors. J Biol Eng 4: 1-14. |
| [29] | Kuimova MK (2012) Mapping viscosity in cells using molecular rotors. Phys Chem Chem Phys 14: 12671-12686. |
| [30] | Thompson AJ, Herling TW, Kubankova M, et al. (2015) Molecular rotors provide insights into microscopic structural changes during protein aggregation. J Phys Chem B 119: 10170-10179. |
| [31] | Vus K, Trusova V, Gorbenko G, et al. (2015) Thioflavin T derivatives for the characterization of insulin and lysozyme amyloid fibrils in vitro: Fluorescence and quantum-chemical studies. J Lumin 159: 284-293. |
| [32] | Yu W-T, Wu T-W, Huang C-L, et al. (2016) Protein sensing in living cells by molecular rotor-based fluorescence-switchable chemical probes. Chem Sci 7: 301-307. |
| [33] | Uzhinov BM, Ivanov VL, Melnikov MY (2011) Molecular rotors as luminescence sensors of local viscosity and viscous flow in solutions and organized systems. Russ Chem Rev 80: 1179-1190. |
| [34] | Jin Y-J, Dogra R, Cheong IW, et al. (2015) Fluorescent molecular rotor-in-paraffin waxes for thermometry and biometric identification. ACS Appl Mater Inter 7: 14485-14492. |
| [35] | F?rster T, Hoffman G (1971) Die Viskosit?tsabh?gigkeit der Fluoreszenzquantenausbeuten eininger Farbstoffsysteme. Z Phys Chem 75: 63-76. |
| [36] | Sutharsan J, Lichlyter D, Wright NE, et al. (2010) Molecular rotors: synthesis and evaluation as viscosity sensors. Tetrahedron 66: 2582-2588. |
| [37] | Sinkeldam RW, Wheat AJ, Boyaci H, et al. (2011) Emissive nucleosides as molecular rotors. Chemphyschem 12: 567-570. |
| [38] | Haidekker MA, Brady TP, Lichlyter D, et al. (2005) Effects of solvent polarity and solvent viscosity on the fluorescent properties of molecular rotors and related probes. Bioorg Chem 33: 415-425. |
| [39] | Ferri G, Nucara L, Biver T, et al. (2016) Organization of inner cellular components as reported by a viscosity-sensitive fluorescent Bodipy probe suitable for phasor approach to FLIM. Biophys Chem 208: 17-25. |
| [40] | Peng X, Yang Z, Wang J, et al. (2011) Fluorescence ratiometry and fluorescence lifetime imaging: Using a single molecular sensor for dual mode imaging of cellular viscosity. J Am Chem Soc 133: 6626-6635. |
| [41] | Wu Y, Stefl M, Olzynska A, et al. (2013) Molecular rheometry: direct determination of viscosity in L-o and L-d lipid phases via fluorescence lifetime imaging. Phys Chem Chem Phys 15: 14986-14993. |
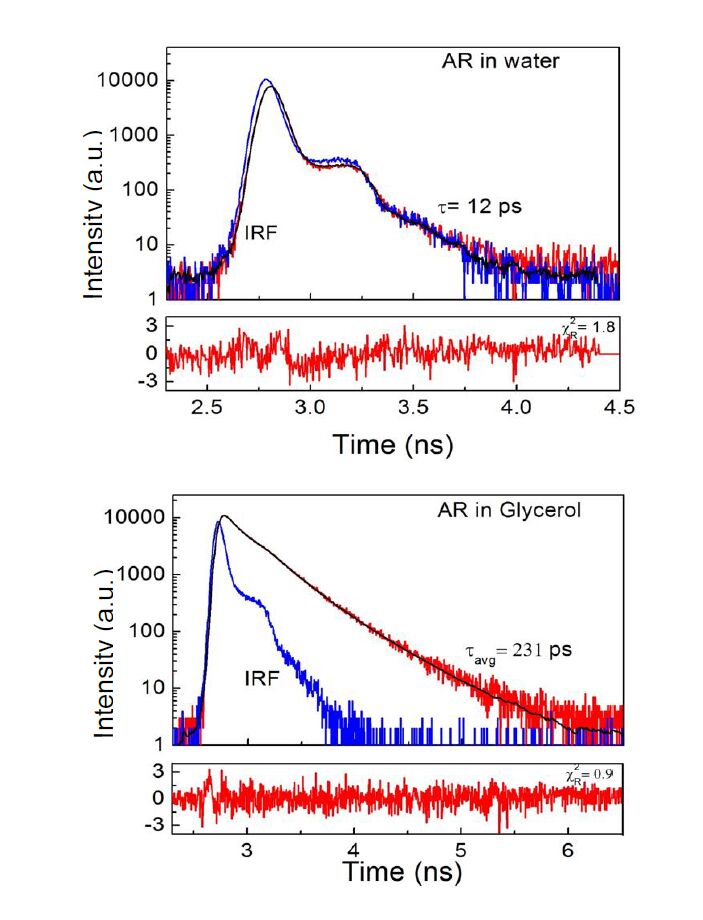
| [42] | Chib R, Shah S, Gryczynski Z, et al. (2016) Standard reference for instrument response function in fluorescence lifetime measurements in visible and near infrared. Meas Science Technol 27: 027001. |
| [43] | Akers W, Haidekker MA (2004) A molecular rotor as viscosity sensor in aqueous colloid solutions. J Biomech Eng T Asme 126: 340-345. |
| [44] | Draganski A (2014) A systematic study of phosphorescent probes in cryosolvents, amorphous solids and proteins. [Ph.D. Dissertation] Rutgers, the State University of New Jersey. |
| [45] | Waxman SM (2016) Synthetic Food Colors: Photophysical Characterization and Potential Uses. [George H. Cook Honors Thesis] Rutgers, The State University of New Jersey. |
| [46] | Moreira PF, Giestas L, Yihwa C, et al. (2003) Ground- and excited-state proton transfer in anthocyanins: From weak acids to superphotoacids. J Phys Chem A 107: 4203-4210. |
| [47] | Castaneda-Ovando A, Pacheco-Hernandez MD, Paez-Hernandez ME, et al. (2009) Chemical studies of anthocyanins: A review. Food Chem 113: 859-871. |
| [48] | Liu CJ, Furusawa Y, Hayashi K (2013) Development of a fluorescent imaging sensor for the detection of human body sweat odor. Sensor Actuat B-Chem 183: 117-123. |
| [49] | Moorthy JN, Shevchenko T, Magon A, et al. (1998) Paper acidity estimation: Application of pH-dependent fluorescence probes. J Photochem Photobiol A 113: 189-195. |
| [50] | Pedros J, Porcar I, Gomez CM, et al. (1997) Interaction of quinine with negatively charged lipid vesicles studied by fluorescence spectroscopy - Influence of the pH. Spectrochim Acta A 53: 421-431. |
| [51] | Slavik J (1994) Fluorescent Probes in Cellular and Molecular Biology. Boca Raton, FL: CRC Press. |
| [52] | Mermelstein NH (2009) Measuring moisture content and water activity. Food Technol 63: 64-68. |
| [53] | Mermelstein NH (2011) Instruments for moisture and water activity determination. Food Technol 65: 68-72. |
| [54] | Jan AT, Kamli MR, Murtaza I, et al. (2010) Dietary flavonoid quercetin and associated health benefits - AnOverview. Food Rev Int 26: 302-317. |
| [55] | Pietta PG (2000) Flavonoids as antioxidants. J Nat Prod 63: 1035-1042. |
| [56] | Guharay J, Dennison SM, Sengupta PK (1999) Influence of different environments on the excited-state proton transfer and dual fluorescence of fisetin. Spectrochim Acta A 55: 1091-1099. |
| [57] | Demchenko AP, Klymchenko AS, Pivovarenko VG, et al. (2003) Multiparametric color-changing fluorescence probes. J Fluoresc 13: 291-295. |
| [58] | Klymchenko AS, Kenfack C, Duportail G, et al. (2007) Effects of polar protic solvents on dual emissions of 3-hydroxychromones. J Chem Sci 119: 83-89. |
| [59] | Tomin VI (2011) Spectral properties of anionic form of 3-hydroxyflavone. Opt Spectrosc 110: 550-556. |
| [60] | Sytnik A, Gormin D, Kasha M (1994) Interplay between excited-state intramolecular proton transfer and charge transfer in flavonols and their use as protein-binding site fluorescence probes. Proc Nat Acad Sci USA 91: 11968-11972. |
| [61] | Ludescher RD, Shah NK, McCaul CP, et al. (2001) optical luminescence measurements of molecular mobility in amorphous solid foods. Food Hydrocolloid 15: 331-339. |
| [62] | Liang J, Wang S, Ludescher RD (2015) Effect of additives on physicochemical properties in amorphous starch matrices. Food Chem 171: 298-305. |
| [63] | Tiwari R, Ludescher RD (2013) Effect of temperature on molecular mobility, oxygen permeability, and dynamic site heterogeneity in amorphous alpha-lactalbumin films. Food Hydrocolloid 31: 357-364. |
| [64] | Joye IJ, Draganski A, Delcour JA, et al. (2012) Monitoring molecular oxygen depletion in wheat flour dough using Erythrosin B phosphorescence: A biophysical approach. Food Biophys 7: 138-144. |
| [65] | You YM, Ludescher RD (2009) Effect of xanthan on the molecular mobility of amorphous sucrose detected by erythrosin B phosphorescence. J Agr Food Food Chem 57: 709-716. |
| [66] | Tiwari R, Ludescher RD (2012) Molecular mobility in a homologous series of amorphous solid glucose oligomers. Food Chem 132: 1814-1821. |
| [67] | Tiwari RS, Ludescher RD (2010) Vanillin phosphorescence as a probe of molecular mobility in amorphous sucrose. J Fluoresc 20: 125-133. |
| [68] | Kuan Lam S, Namdas E, Lo D (1998) Effects of oxygen and temperature on phosphorescence and delayed fluorescence of erythrosin B trapped in sol-gel silica. J Photochem Photobiol A 118: 25-30. |
| [69] | Chan MA, Lam SK, Lo D (2002) Characterization of erythrosin B-doped sol-gel materials for oxygen sensing in aqueous solutions. J Fluoresc 12: 327-332. |
| [70] | Liang J, Xia QY, Wang SM, et al. (2015) Influence of glycerol on the molecular mobility, oxygen permeability and microstructure of amorphous zein films. Food Hydrocolloid 44: 94-100. |
| [71] | You YM, Ludescher RD (2008) The effect of salts on molecular mobility in amorphous sucrose monitored by erythrosin B phosphorescence. Carbohyd Res 343: 2641-2649. |
| [72] | You YM, Ludescher RD (2008) Effect of gelatin on molecular mobility in amorphous sucrose detected by erythrosin B phosphorescence. Carbohyd Res 343: 2657-2666. |
| [73] | Schlyer BD, Schauerte JA, Steel DG, et al. (1994) Time-resolved room temperature protein phosphorescence: nonexponential decay from single emitting tryptophans. Biophys J 67: 1192-1202. |
| [74] | Steel DG, Subramaniam V, Gafni A (1996) Time-resolved tryptophan phosphorescence spectroscopy: A sensitive probe of protein folding and structure. IEEE J Sel Top Quantum 2: 1107-1114. |
| [75] | Penzkofer A, Tyagi A, Slyusareva E, et al. (2010) Phosphorescence and delayed fluorescence properties of fluorone dyes in bio-related films. Chem Phys 378: 58-65. |
| [76] | Wang Y (2016) A study of luminescent properties of riboflavin and its potential applications as a luminescent probe in food science. [PhD Dissertation] Rutgers, The State University of New Jersey. |
| [77] | Moore WM, McDaniels JC, Hen JA (1977) The photochemistry of riboflavin—vi. The photophysical properties of isoalloxazines. Photochem Photobiol 25: 505-512. |
| [78] | Grodowski MS, Veyret B, Weiss K (1977) Photochemistry of flavins. ii. Photophysical properties of alloxazines and isoalloxazines. Photochem Photobiol 26: 341-352. |
| [79] | Ahmad I, vaid F (2006) Photochemistry of flavins in aqueous and organic solvents. Photochem Photobiol 2: 13-40. |
| [80] | Gandia-Herrero F, Escribano J, Garcia-Carmona F (2005) Betaxanthins as pigments responsible for visible fluorescence in flowers. Planta 222: 586-593. |
| [81] | Gandia-Herrero F, Garcia-Carmona F, Escribano J (2005) A novel method using high-performance liquid chromatography with fluorescence detection for the determination of betaxanthins. J Chromatogr A 1078: 83-89. |
| [82] | Wang SY, Le AN, Corradini MG, et al. (2015) Flavonols as luminescent probes of water activity in foods and pharmaceuticals. Biophys J 108: 492a-492a. |
| [83] | Li YX, Zou XB, Huang XW, et al. (2015) A new room temperature gas sensor based on pigment-sensitized TiO2 thin film for amines determination. Biosens Bioelectron 67: 35-41. |
| [84] | Kuswandi B, Jayus, Larasati TS, et al. (2012) Real-time monitoring of shrimp spoilage using on-package sticker sensor based on natural dye of curcumin. Food Anal Methods 5: 881-889. |
| [85] | Priyadarsini KI (2009) Photophysics, Photochem Photobiol of curcumin: Studies from organic solutions, bio-mimetics and living cells. J Photoch Photobiol C 10: 81-95. |
| [86] | Draganski AR, Tiwari RS, Sundaresan KV, et al. (2010) Photophysical probes of the amorphous solid state of proteins. Food Biophys 5: 337-345. |
| [87] | Narsaiah K, Jha SN, Brardwai R, et al. (2012) Optical biosensors for food quality and safety assurance—a review. J Food Sci Technol 49: 383-406. |
| [88] | Mello LD, Kubota LT (2002) Review of the use of biosensors as analytical tools in the food and drink industries. Food Chem 77: 237-256. |
| [89] | Leung C-H, Chan DS-H, Man BY-W, et al. (2011) Simple and Convenient G-Quadruplex-Based Turn-On Fluorescence Assay for 3′ → 5′ Exonuclease Activity. Anal Chem 83: 463-466. |
Figures(8) / Tables(3)

G.Corradini Maria, LaviniaWang Yan, Le An, M.Waxman Sarah, Zelent Bogumil, Chib Rahul, Gryczynski Ignacy, D.Ludescher Richard. Identifying and selecting edible luminescent probes as sensors of food quality[J]. AIMS Biophysics, 2016, 3(2): 319-339. doi: 10.3934/biophy.2016.2.319







 DownLoad:
DownLoad: