Citation: David Kirchenbuechler, Yael Mutsafi, Ben Horowitz, Smadar Levin-Zaidman, Deborah Fass, Sharon G. Wolf, Michael Elbaum. Cryo-STEM Tomography of Intact Vitrified Fibroblasts[J]. AIMS Biophysics, 2015, 2(3): 259-273. doi: 10.3934/biophy.2015.3.259

| [1] |
Bai X, McMullan G, Scheres SHW (2015) How cryo-EM is revolutionizing structural biology. Trends Biochem Sci 40: 49-57. doi: 10.1016/j.tibs.2014.10.005

|
| [2] | Kühlbrandt W (2014) Cryo-EM enters a new era. eLife 3: e03678. |
| [3] |
Briegel A, Dias DP, Li Z, et al. (2006) Multiple large filament bundles observed in Caulobactercrescentus by electron cryotomography. Mol Microbiol 62: 5-14. doi: 10.1111/j.1365-2958.2006.05355.x
|
| [4] | Henderson GP, Gan L, Jensen GJ (2007) 3-D ultrastructure of O. tauri: electron cryotomography of an entire eukaryotic cell. PloS One 2: e749. |
| [5] | Danev R, Kanamaru S, Marko M, et al. (2010) Zernike phase contrast cryo-electron tomography. J Struct Biol 171: 174-181. |
| [6] |
Murata K, Liu X, Danev R, et al. (2010) Zernike Phase Contrast Cryo-Electron Microscopy and Tomography for Structure Determination at Nanometer and Subnanometer Resolutions. Structure 18: 903-912. doi: 10.1016/j.str.2010.06.006
|
| [7] | Guerrero-Ferreira RC, Wright ER (2014) Zernike phase contrast cryo-electron tomography of whole bacterial cells. J Struct Biol 185: 129-133. |
| [8] |
Danev R, Buijsse B, Khoshouei M, et al. (2014) Volta potential phase plate for in-focus phase contrast transmission electron microscopy. Proc Natl Acad Sci U S A 111: 15635-15640. doi: 10.1073/pnas.1418377111
|
| [9] | Briggs JAG (2013) Structural biology in situ--the potential of subtomogram averaging. Curr Opin Struct Biol 23: 261-267. |
| [10] |
Marko M, Hsieh C, Schalek R, et al. (2007) Focused-ion-beam thinning of frozen-hydrated biological specimens for cryo-electron microscopy. Nat Methods 4: 215-217. doi: 10.1038/nmeth1014
|
| [11] | Hsieh C, Schmelzer T, Kishchenko G, et al. (2014) Practical workflow for cryo focused-ion-beam milling of tissues and cells for cryo-TEM tomography. J Struct Biol 185: 32-41. |
| [12] | Rigort A, Bäuerlein FJB, Villa E, et al. (2012) Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proc Natl Acad Sci U S A 109: 4449-4454. |
| [13] |
Kapishnikov S, Weiner A, Shimoni E, et al. (2012) Oriented nucleation of hemozoin at the digestive vacuole membrane in Plasmodium falciparum. Proc Natl Acad Sci U S A 109: 11188-11193. doi: 10.1073/pnas.1118120109
|
| [14] |
Müller WG, Heymann JB, Nagashima K, et al. (2012) Towards an atlas of mammalian cell ultrastructure by cryo soft X-ray tomography. J Struct Biol 177: 179-192. doi: 10.1016/j.jsb.2011.11.025
|
| [15] |
Carzaniga R, Domart M-C, Collinson LM, et al. (2014) Cryo-soft X-ray tomography: a journey into the world of the native-state cell. Protoplasma 251: 449-458. doi: 10.1007/s00709-013-0583-y
|
| [16] | Cruz-Adalia A, Ramirez-Santiago G, Calabia-Linares C, et al. (2014) T cells kill bacteria captured by transinfection from dendritic cells and confer protection in mice. Cell Host Microbe 15: 611-622. |
| [17] | Do M, Isaacson SA, McDermott G, et al. (2015) Imaging and characterizing cells using tomography. Arch Biochem Biophys. |
| [18] |
Schertel A, Snaidero N, Han H-M, et al. (2013) Cryo FIB-SEM: Volume imaging of cellular ultrastructure in native frozen specimens. J Struct Biol 184: 355-360. doi: 10.1016/j.jsb.2013.09.024
|
| [19] | Wolf SG, Houben L, Elbaum M (2014) Cryo-scanning transmission electron tomography of vitrified cells. Nat Methods 11: 423-428. |
| [20] | Yakushevska AE, Lebbink MN, Geerts WJC, et al. (2007) STEM tomography in cell biology. J Struct Biol 159: 381-391. |
| [21] | Aoyama K, Takagi T, Hirase A, et al. (2008) STEM tomography for thick biological specimens. Ultramicroscopy 109: 70-80. |
| [22] |
Hohmann-Marriott MF, Sousa AA, Azari AA, et al. (2009) Nanoscale 3D cellular imaging by axial scanning transmission electron tomography. Nat Methods 6: 729-731. doi: 10.1038/nmeth.1367
|
| [23] |
Kremer JR, Mastronarde DN, McIntosh JR (1996) Computer visualization of three-dimensional image data using IMOD. J Struct Biol 116: 71-76. doi: 10.1006/jsbi.1996.0013
|
| [24] |
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9: 671-675. doi: 10.1038/nmeth.2089
|
| [25] |
Heymann JB, Belnap DM (2007) Bsoft: image processing and molecular modeling for electron microscopy. J Struct Biol 157: 3-18. doi: 10.1016/j.jsb.2006.06.006
|
| [26] | Pettersen EF, Goddard TD, Huang CC, et al. (2004) UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 25: 1605-1612. |
| [27] |
Jensen GJ, Kornberg RD (2000) Defocus-gradient corrected back-projection. Ultramicroscopy 84: 57-64. doi: 10.1016/S0304-3991(00)00005-X
|
| [28] |
Philippsen A, Engel H-A, Engel A (2007) The contrast-imaging function for tilted specimens. Ultramicroscopy 107: 202-212. doi: 10.1016/j.ultramic.2006.07.010
|
| [29] |
Zanetti G, Riches JD, Fuller SD, et al. (2009) Contrast transfer function correction applied to cryo-electron tomography and sub-tomogram averaging. J Struct Biol 168: 305-312. doi: 10.1016/j.jsb.2009.08.002
|
| [30] |
Voortman LM, Franken EM, van Vliet LJ, et al. (2012) Fast, spatially varying CTF correction in TEM. Ultramicroscopy 118: 26-34. doi: 10.1016/j.ultramic.2012.05.002
|
| [31] |
Rez P (2003) Comparison of phase contrast transmission electron microscopy with optimized scanning transmission annular dark field imaging for protein imaging. Ultramicroscopy 96: 117-124. doi: 10.1016/S0304-3991(02)00436-9
|
| [32] |
Engel A (2009) Chapter 9 Scanning Transmission Electron Microscopy: Biological Applications. Adv Imag Elect Phys 159: 357-386. doi: 10.1016/S1076-5670(09)59009-X
|
| [33] |
Reichelt R, Holzenburg A, Buhle EL, et al. (1990) Correlation between structure and mass distribution of the nuclear pore complex and of distinct pore complex components. J Cell Biol 110: 883-894. doi: 10.1083/jcb.110.4.883
|
| [34] |
Trachtenberg S, Leonard KR, Tichelaar W (1992) Radial mass density functions of vitrified helical specimens determined by scanning transmission electron microscopy: their potential use as substitutes for equatorial data. Ultramicroscopy 45: 307-321. doi: 10.1016/0304-3991(92)90142-7
|
| [35] |
Angert I, Majorovits E, Schröder RR (2000) Zero-loss image formation and modified contrast transfer theory in EFTEM. Ultramicroscopy 81: 203–222. doi: 10.1016/S0304-3991(99)00190-4
|
| [36] | Grimm R, Typke D, Bärmann M, et al. (1996) Determination of the inelastic mean free path in ice by examination of tilted vesicles and automated most probable loss imaging. Ultramicroscopy 63: 169–179. |
| [37] |
Baudoin JP1, Jinschek JR, Boothroyd CB, et al. (2013) Chromatic aberration-corrected tilt series transmission electron microscopy of nanoparticles in a whole mount macrophage cell. Microsc Microanal 19: 814-820. doi: 10.1017/S1431927613001475
|
| [38] |
Henderson R (1995) The potential and limitations of neutrons, electrons and X-rays for atomic resolution microscopy of unstained biological molecules. Q Rev Biophys 28: 171-193. doi: 10.1017/S003358350000305X
|
| [39] |
Pennycook SJ (1989) Z-contrast stem for materials science. Ultramicroscopy 30: 58-69. doi: 10.1016/0304-3991(89)90173-3
|
| [40] | Carlemalm E, Kellenberger E (1982) The reproducible observation of unstained embedded cellular material in thin sections: visualisation of an integral membrane protein by a new mode of imaging for STEM. EMBO J 1: 63-67. |
| [41] | Krivanek OL, Chisholm MF, Nicolosi V, et al. (2010) Atom-by-atom structural and chemical analysis by annular dark-field electron microscopy. Nature 464: 571-574. |
| [42] | Wolf SG, Rez P, Elbaum M (2015) Phosphorus detection in vitrified bacteria by cryo-STEM annular dark field analysis. J Microscopy doi/10.1111/jmi.12289 (e-print, in press). |
| [43] |
Hagen C, Guttmann P, Klupp B, et al. (2012) Correlative VIS-fluorescence and soft X-ray cryo-microscopy/tomography of adherent cells. J Struct Biol 177: 193-201. doi: 10.1016/j.jsb.2011.12.012
|
| [44] |
Duke EMH, Razi M, Weston A, et al. (2014) Imaging endosomes and autophagosomes in whole mammalian cells using correlative cryo-fluorescence and cryo-soft X-ray microscopy (cryo-CLXM). Ultramicroscopy 143:77-87. doi: 10.1016/j.ultramic.2013.10.006
|
| [45] |
Kapishnikov S, Weiner A, Shimoni E, et al. (2013) Digestive Vacuole Membrane in Plasmodium falciparum-Infected Erythrocytes: Relevance to Templated Nucleation of Hemozoin. Langmuir 29: 14595-14602. doi: 10.1021/la402545c
|
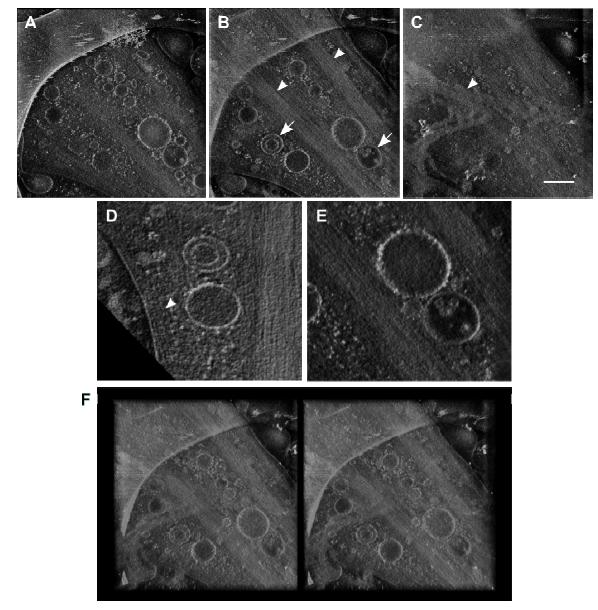
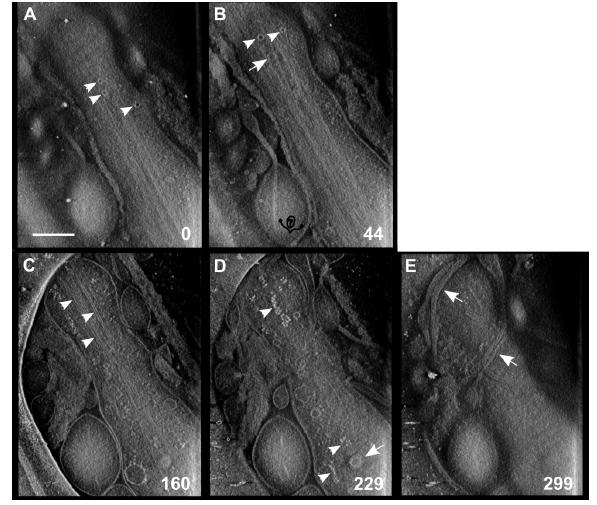

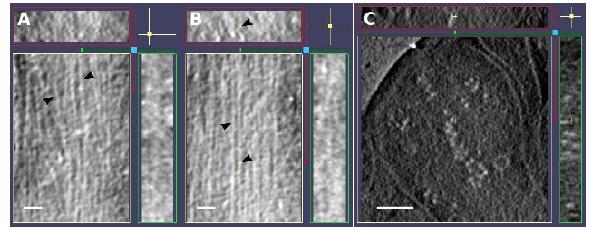
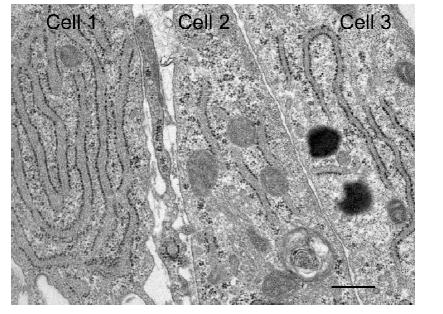
Figures(6)

David Kirchenbuechler, Yael Mutsafi, Ben Horowitz, Smadar Levin-Zaidman, Deborah Fass, Sharon G. Wolf, Michael Elbaum. Cryo-STEM Tomography of Intact Vitrified Fibroblasts[J]. AIMS Biophysics, 2015, 2(3): 259-273. doi: 10.3934/biophy.2015.3.259







 DownLoad:
DownLoad: