Citation: Albert Henry Ntarmah, Yusheng Kong, Michael Kobina Gyan. Banking system stability and economic sustainability: A panel data analysis of the effect of banking system stability on sustainability of some selected developing countries[J]. Quantitative Finance and Economics, 2019, 3(4): 709-738. doi: 10.3934/QFE.2019.4.709

| [1] |
Al-Moulani A, Constantinos A (2017) Banking sector depth and economic growth nexus: a comparative study between the natural resource-based and the rest of the world's economies, International. Rev Appl Econ 31: 625-650. DOI:10.1080/02692171.2017.1299115 doi: 10.1080/02692171.2017.1299115

|
| [2] | Alexander K (2014) Stability and Sustainability in Banking Reform: are environmental risks missing in Basel III. CISL & UNEPFI: Cambridge and Geneva. |
| [3] |
Aluko OA, Ajayi MA (2018) Determinants of banking sector development: Evidence from Sub-Saharan African countries. Borsa Istanbul Rev 18: 122-139. doi: 10.1016/j.bir.2017.11.002
|
| [4] | Arcand JL, Berkes E, Panizza U (2012) Too much finance? IMF Working Paper, No. 12/161, Washington DC: IMF. |
| [5] | Azeez BA, Oke MO (2012) A Time Series Analysis on the Effect of Banking Reforms on Nigeria's Economic Growth. Int J Econ Res 3: 26-37. |
| [6] | Baltagi BH (2005) Econometric Analysis of Data Panel, England: John Wiley & Sons Ltd. |
| [7] | Bolt K, Matete M, Clemens M (2002) Manual for calculating adjusted net savings. Environ Department World Bank, 1-23. |
| [8] | Bryman A, Bell E (2015) Business research methods, Oxford University Press, USA. |
| [9] |
Demetriades PO, Rousseau PL (2016) The changing face of financial development. Econ Lett 141: 87-90. doi: 10.1016/j.econlet.2016.02.009
|
| [10] |
Charfeddine L, Ben Khediri K (2016) Financial development and environmental quality in UAE: Cointegration with structural breaks. Renew Sust Energy Rev 55: 1322-1335. doi: 10.1016/j.rser.2015.07.059
|
| [11] | Demirgüç-Kunt A, Detragiache E (1999) The determinants of banking crises: evidence from industrial and developing countries, The World Bank. |
| [12] | Demirgüç-Kunt A, Detragiache E (2005) Cross-country empirical studies of systemic bank distress: a survey, The World Bank. |
| [13] |
Ferreira S, Vincent JR (2005) Genuine savings: leading indicator of sustainable development?. Econ Dev Cult Change 53: 737-754. doi: 10.1086/426834
|
| [14] |
Ferreira S, Hamilton K, Vincent JR (2008) Comprehensive wealth and future consumption: accounting for population growth. World Bank Econ Rev 22: 233-248. doi: 10.1093/wber/lhn008
|
| [15] | Genberg H (2017) Global shocks and risk to financial stability in Asia (No. 709). ADBI Working Paper Series. |
| [16] | Hamilton K (2006) Where is the wealth of nations?: measuring capital for the 21st century, World Bank Publications. |
| [17] |
Hanley N, Dupuy L, McLaughlin E (2015) Genuine savings and sustainability. J Econ Surv 29: 779-806. doi: 10.1111/joes.12120
|
| [18] |
Jacobs LM, Van Rossem R (2014) The BRIC Phantom: A comparative analysis of the BRICs as a category of rising powers. J Policy Model 36: S47-S66. doi: 10.1016/j.jpolmod.2013.10.008
|
| [19] |
Jayakumar M, Pradhan RP, Dash S, et al. (2018) Banking competition, banking stability, and economic growth: Are feedback effects at work? J Econ Bus 96: 15-41. doi: 10.1016/j.jeconbus.2017.12.004
|
| [20] | Jiang B (2014) Financial stability of banking system in China, Doctoral dissertation, Nottingham Trent University. |
| [21] |
Jokipii T, Monnin P (2013) The impact of banking sector stability on the real economy. J Int Money Financ 32: 1-16. doi: 10.1016/j.jimonfin.2012.02.008
|
| [22] | Jung JY (2008) Regional financial cooperation in Asia: challenges and path to development. Press & Commun CH 4002 Basel Switzerland, 120. |
| [23] |
Kaminsky GL, Reinhart CM (1999) The twin crises: the causes of banking and balance-of-payments problems. Am Econ Rev 89: 473-500. doi: 10.1257/aer.89.3.473
|
| [24] |
Kregel J (2009) The global crisis and the implications for developing countries and the BRICs: Is the "B" really justified? Braz J Polit Econ 29: 341-356. doi: 10.1590/S0101-31572009000400002
|
| [25] | Moundigbaye M, Rea WS, Reed WR (2017) Which panel data estimator should I use?: A corrigendum and extension. Economics Discussion Papers, No 2017-58. Kiel Inst World Econ 2. |
| [26] | Mhadhbi K (2014) Financial development and economic growth: A dynamic panel data analysis. Int J Econ Financ Manage 2: 48-58. |
| [27] | Murphy P, Waygood S, Wilson G (2017) Ideas for Action for a Long-Term and Sustainable Financial System. Bus Sustainable Dev Comm. |
| [28] |
Narayan PK, Narayan S (2013) The short-run relationship between the financial system and economic growth: New evidence from regional panels. Int Rev Financ Anal 29: 70-78. doi: 10.1016/j.irfa.2013.03.012
|
| [29] | Pardi F, Salleh AM, Nawi AS (2015) Determinants of Sustainable Development in Malaysia: A VECM Approach of Short-Run and Long-Run Relationships. Am J Econ 5: 269-277. |
| [30] | Pazarbasioglu C, Johnsen G, Hilbers MPLC, et al. (2005) Assessing and managing rapid credit growth and the role of supervisory and prudential policies (No. 5-151), International Monetary Fund. |
| [31] |
Pearce DW, Atkinson GD (1993) Capital theory and the measurement of sustainable development: an indicator of "weak" sustainability. Ecol Econ 8: 103. doi: 10.1016/0921-8009(93)90039-9
|
| [32] |
Pillarisetti JR (2005) The World Bank's 'genuine savings' measure and sustainability. Ecol Econ 55: 599-609. doi: 10.1016/j.ecolecon.2005.01.018
|
| [33] |
Pradhan RP, Arvin MB, Nair M, et al. (2019) Inter-linkages between competition and stabilisation policies in the banking sector and stock market development in Europe. Appl Econ 51: 4313-4324. doi: 10.1080/00036846.2019.1591603
|
| [34] |
Rahman MM, Ashraf BN, Zheng C, et al. (2017) Impact of Cost Efficiency on Bank Capital and the Cost of Financial Intermediation: Evidence from BRICS Countries. Int J Financ Stud 5: 32. doi: 10.3390/ijfs5040032
|
| [35] | Schmidt-Traub G, Shah A (2015) Investment needs to achieve the Sustainable Development Goals. Paris New York Sustainable Dev Solutions Network. |
| [36] | Sen S (2016) The BRICS Initiatives Towards a New Financial Architecture: An Assessment with Some Proposals (No. 205). RIS Discussion Paper. |
| [37] |
Solow RM (1956) A Contribution to the Theory of Economic Growth. Q J Econ 70: 65-94. doi: 10.2307/1884513
|
| [38] | Stewart R, Chowdhury M (2019) Does Bank Stability Promote Economic Resilience?: Evidence From Panel Data. Evidence From Panel Data. |
| [39] | TheGlobalEconomy.com (2019) Business and economic data for 200 countries. Available from https://www.theglobaleconomy.com/indicators_list.php. |
| [40] |
Tongurai J, Vithessonthi C (2018) The impact of the banking sector on economic structure and growth. Int Rev Financ Anal 56: 193-207. doi: 10.1016/j.irfa.2018.01.002
|
| [41] |
Tripathy R (2019) Doe measure of financial development matter for economic growth in India? Quant Financ Econ 3: 508-525. doi: 10.3934/QFE.2019.3.508
|
| [42] | World Bank (2012) Contribution to Beyond GDP, "Virtual Indicator Expo", Washington DC: World Bank. |
| [43] | World Bank (2019) Economic research data. Retrieved from FRED, Federal Reserve Bank of St. Louis. [dataset for banking system stability] Available from: https://fred.stlouisfed.org/series. |
| [44] | World Bank Group (2019) World Development Indicators 2019, Washington DC: World Bank. [dataset for economic sustainability and other determinants]. |
| [45] | Wyman O (2015) Post-Crisis Changes in the Stability of the US Banking System Evidence from US Bank Holding Companies from 2004 to 2014, NY 10036: New York. |
| [46] | Yaffee R (2003) A primer for panel data analysis. Connect: Information Technology at NYU, 1-11. |
| [47] | Younsi M, Bechtini M (2018) Economic Growth, Financial Development and Income Inequality in BRICS Countries: Evidence from Panel Granger Causality Tests. Available from: https://mpra.ub.uni-muenchen.de/85182/. |
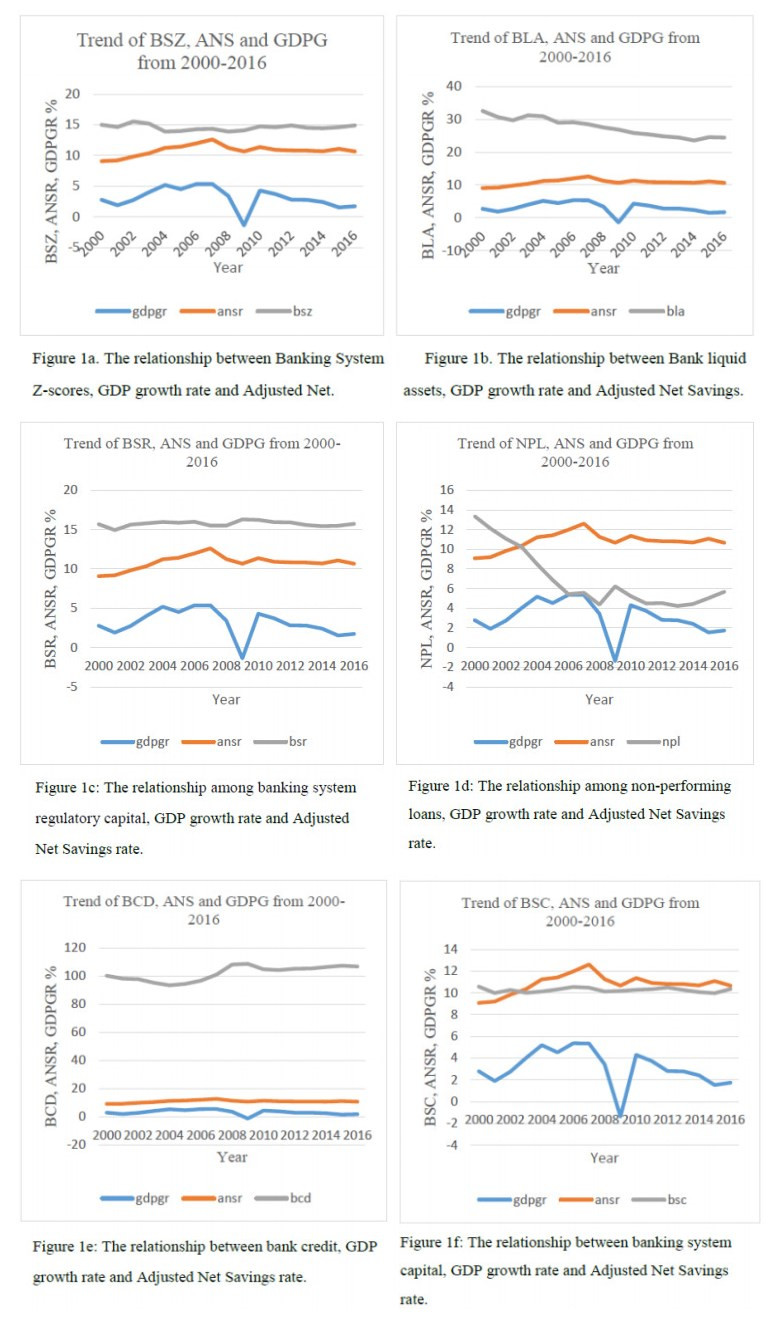
Figures(1) / Tables(10)

Albert Henry Ntarmah, Yusheng Kong, Michael Kobina Gyan. Banking system stability and economic sustainability: A panel data analysis of the effect of banking system stability on sustainability of some selected developing countries[J]. Quantitative Finance and Economics, 2019, 3(4): 709-738. doi: 10.3934/QFE.2019.4.709







 DownLoad:
DownLoad: