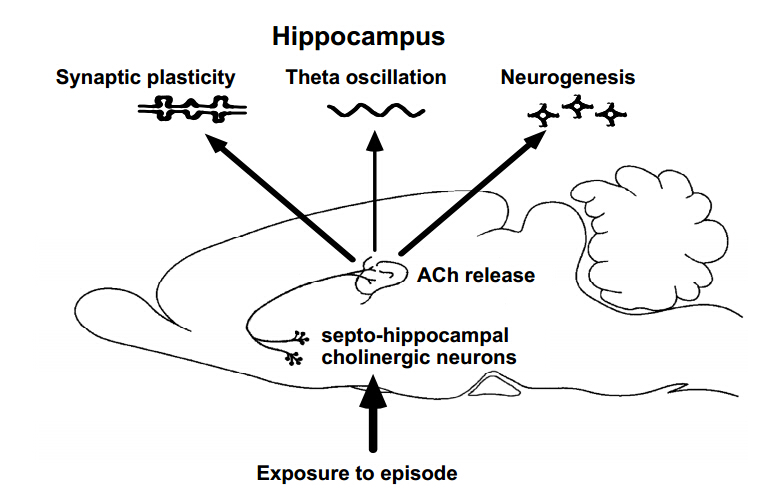
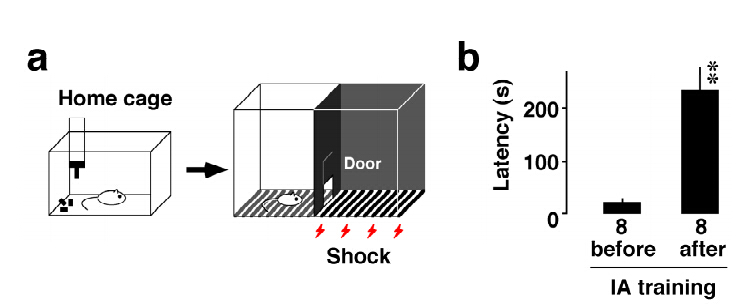
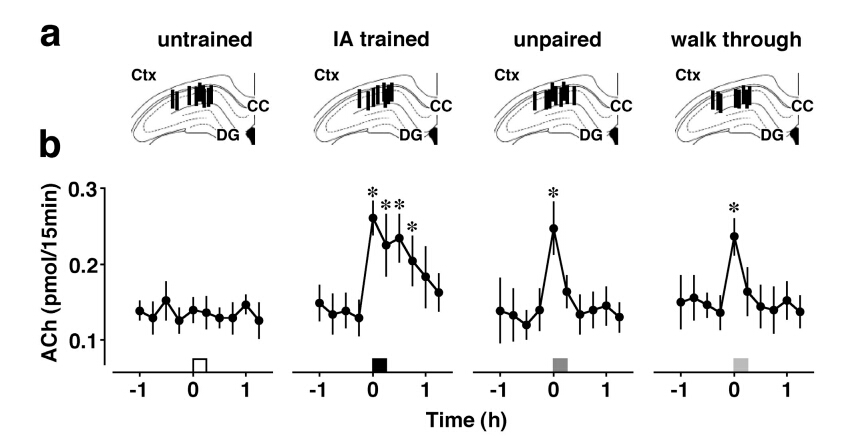
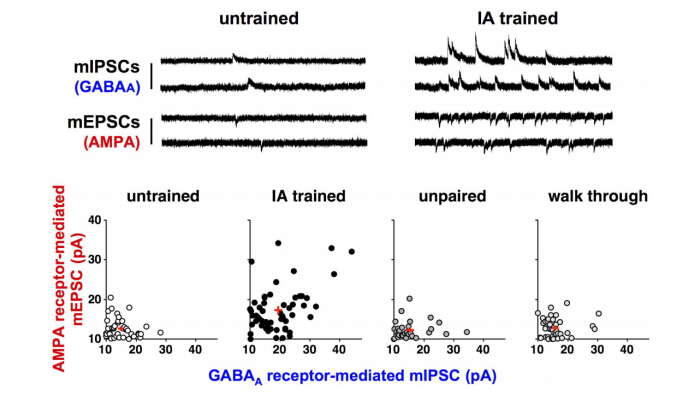
Although the hippocampus is processing temporal and spatial information in particular context, the encoding rule creating memory is completely unknown. To examine the mechanism, we trained rats on an inhibitory avoidance (IA) task, a hippocampus-dependent rapid one-trial contextual learning paradigm. By combining Herpes virus-mediated in vivo gene delivery with in vitro patch-clamp recordings, I reported contextual learning drives GluR1-containing AMPA receptors into CA3-CA1 synapses. The molecular event is required for contextual memory, since bilateral expression of delivery blocker in CA1 successfully blocked IA learning. Moreover, I found a logarithmic correlation between the number of delivery blocking cells and learning performance. Considering that one all-or-none device can process 1-bit of data per clock (Nobert Wiener 1961), the logarithmic correlation may provides evidence that CA1 neurons transmit essential data of contextual information. Further, I recently reported critical role of acetylcholine as an intrinsic trigger of learning-dependent synaptic plasticity. IA training induced ACh release in CA1 that strengthened not only AMPA receptor-mediated excitatory synapses, but also GABAA receptor-mediated inhibitory synapses on each CA1 neuron. More importantly, IA-trained rats showed individually different excitatory and inhibitory synaptic inputs with wide variation on each CA1 neuron. Here I propose a new hypothesis that the diversity of synaptic inputs on CA1 neurons may depict cell-specific outputs processing experienced episodes after training.
Citation: Dai Mitsushima. Contextual Learning Requires Functional Diversity at Excitatory and Inhibitory Synapses onto CA1 Pyramidal Neurons[J]. AIMS Neuroscience, 2015, 2(1): 7-17. doi: 10.3934/Neuroscience.2015.1.7
Although the hippocampus is processing temporal and spatial information in particular context, the encoding rule creating memory is completely unknown. To examine the mechanism, we trained rats on an inhibitory avoidance (IA) task, a hippocampus-dependent rapid one-trial contextual learning paradigm. By combining Herpes virus-mediated in vivo gene delivery with in vitro patch-clamp recordings, I reported contextual learning drives GluR1-containing AMPA receptors into CA3-CA1 synapses. The molecular event is required for contextual memory, since bilateral expression of delivery blocker in CA1 successfully blocked IA learning. Moreover, I found a logarithmic correlation between the number of delivery blocking cells and learning performance. Considering that one all-or-none device can process 1-bit of data per clock (Nobert Wiener 1961), the logarithmic correlation may provides evidence that CA1 neurons transmit essential data of contextual information. Further, I recently reported critical role of acetylcholine as an intrinsic trigger of learning-dependent synaptic plasticity. IA training induced ACh release in CA1 that strengthened not only AMPA receptor-mediated excitatory synapses, but also GABAA receptor-mediated inhibitory synapses on each CA1 neuron. More importantly, IA-trained rats showed individually different excitatory and inhibitory synaptic inputs with wide variation on each CA1 neuron. Here I propose a new hypothesis that the diversity of synaptic inputs on CA1 neurons may depict cell-specific outputs processing experienced episodes after training.

| [1] |
Scoville WB, Milner B (1957) Loss of recent memory after bilateral hippocampal lesions. J Neurology, Neurosurgery and Psychiatry 20: 11-21. doi: 10.1136/jnnp.20.1.11

|
| [2] |
Wills TJ, Cacucci F, Burgess N, et al. (2010) Development of the hippocampal cognitive map in preweanling rats. Science 328: 1573-1576. doi: 10.1126/science.1188224
|
| [3] | Mitsushima D, Takase K, Funabashi T, et al. (2009) Gonadal steroids maintain 24-h acetylcholine release in the hippocampus: organizational and activational effects in behaving rats. J Neurosci 29:3808–3815. |
| [4] |
Chen G, Wang LP, Tsien JZ (2009) Neural population-level memory traces in the mouse hippocampus. PLoS ONE 4: e8256. doi: 10.1371/journal.pone.0008256
|
| [5] |
Gelbard-Sagiv H, Mukamel R, Harel M, et al. (2008) Internally generated reactivation of single neurons in human hippocampus during free recall. Science 322: 96-101. doi: 10.1126/science.1164685
|
| [6] | Morris RGM, Anderson E, Lynch GS, et al. (1986) Selective impairment of learning and blockade of long-term potentiation by an N-methyl-D-aspartate receptor antagonist, AP5. Nature319: 774-776. |
| [7] |
Bliss TVP, Làmo T (1973) Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J Physiol 232:331-356. doi: 10.1113/jphysiol.1973.sp010273
|
| [8] |
Whitlock JR, Heynen AJ, Shuler MG, B et al. (2006) Learning induces long-term potentiation in the hippocampus. Science 313: 1093-1097. doi: 10.1126/science.1128134
|
| [9] |
Mitsushima D, Ishihara K, Sano A, et al. (2011) Contextual learning requires synaptic AMPA receptor delivery in the hippocampus. Proc Natl Acad Sci USA 108: 12503-12508. doi: 10.1073/pnas.1104558108
|
| [10] |
Fisahn A, Pike FG, Buhl EH, et al. (1998) Cholinergic induction of networkoscillations at 40Hz in the hippocampus in vitro. Nature 394: 186-189. doi: 10.1038/28179
|
| [11] |
Auerbach JM, Segal M (1996) Muscarinic receptors mediating depression and long-term potentiation in rat hippocampus. J Physiol 492: 479-493. doi: 10.1113/jphysiol.1996.sp021323
|
| [12] | Mitsushima D, Sano A, Takahashi T (2013) A cholinergic trigger drives learning-induced plasticity at hippocampal synapses. Nat Commun 4:2760. |
| [13] |
Okada D, Ozawa F, Inokuchi K (2009) Input-specific spine entry of soma-derived Vesl-1S protein conforms to synaptic tagging. Science 324: 904-909. doi: 10.1126/science.1171498
|
| [14] |
Lesburguères E, Gobbo OL, Alaux-Cantin S, et al. (2011) Early tagging of cortical networks is required for the formation of enduring associative memory. Science 331: 924-928. doi: 10.1126/science.1196164
|
| [15] |
Ragozzino ME, Unick KE, Gold PE (1996) Hippocampal acetylcholine release during memory testing in rats: augmentation by glucose. Proc Natl Acad Sci USA 93: 4693-4698. doi: 10.1073/pnas.93.10.4693
|
| [16] |
Stancampiano R, Cocco S, Cugusi C, et al. (1999) Serotonin and acetylcholine release response in the rat hippocampus during a spatial memory task. Neuroscience 89: 1135-1143. doi: 10.1016/S0306-4522(98)00397-2
|
| [17] |
Hironaka N, Tanaka K, Izaki Y, et al. (2001) Memory-related acetylcholine efflux from the rat prefrontal cortex and hippocampus: a microdialysis study. Brain Res 901: 143-150. doi: 10.1016/S0006-8993(01)02338-1
|
| [18] |
Gold PE (2003) Acetylcholine modulation of neural systems involved in learning and memory. Neurobiol Learn Mem 80: 194-210. doi: 10.1016/j.nlm.2003.07.003
|
| [19] |
Parent MB, Baxter MG (2004) Septohippocampal acetylcholine: involved in but not necessary for learning and memory? Learn Mem 11: 9-20. doi: 10.1101/lm.69104
|
| [20] |
Herrera-Morales W, Mar I, Serrano B, et al. (2007) Activation of hippocampal postsynaptic muscarinic receptors is involved in long-term spatial memory formation. Eur J Neurosci 25:1581-1588. doi: 10.1111/j.1460-9568.2007.05391.x
|
| [21] |
Lee MG, Chrobak JJ, Sik A, et al. (1994) Hippocampal theta activity following selective lesion of the septal cholinergic system. Neuroscience 62: 1033-1047. doi: 10.1016/0306-4522(94)90341-7
|
| [22] | Hyman JM, Wyble BP, Goyal V, et al. (2003) Stimulation in hippocampal region CA1 in behaving rats yields long-term potentiation when delivered to the peak of theta and long-term depression when delivered to the trough. J Neurosci 23: 11725-11731. |
| [23] |
Seeger T, Fedorova I, Zheng F, et al. (2004) M2 muscarinic acetylcholine receptor knock-out mice show deficits in behavioral flexibility, working memory, and hippocampal plasticity. J Neurosci 24: 10117-10127. doi: 10.1523/JNEUROSCI.3581-04.2004
|
| [24] |
Shinoe T, Matsui M, Taketo MM, et al. (2005) Modulation of synaptic plasticity by physiological activation of M1 muscarinic acetylcholine receptors in the mouse hippocampus. J Neurosci 25: 11194-11200. doi: 10.1523/JNEUROSCI.2338-05.2005
|
| [25] |
Cole AE, Nicoll RA (1983) Acetylcholine mediates a slow synaptic potential in hippocampal pyramidal cells. Science 221: 1299-1301. doi: 10.1126/science.6612345
|
| [26] |
Markram H, Segal M (1990) Long-lasting facilitation of excitatory postsynaptic potentials in the rat hippocampus by acetylcholine. J Physiol 427: 381-393. doi: 10.1113/jphysiol.1990.sp018177
|
| [27] |
Widmer H, Ferrigan L, Davies CH, et al. (2006) Evoked slow muscarinic acetylcholinergic synaptic potentials in rat hippocampal interneurons. Hippocampus 16: 617-628. doi: 10.1002/hipo.20191
|
| [28] |
Fernández de Sevilla D, Núñez A, Borde M, et al. (2008) Cholinergic-mediated IP3-receptor activation induces long-lasting synaptic enhancement in CA1 pyramidal neurons. J Neurosci 28:1469-1478. doi: 10.1523/JNEUROSCI.2723-07.2008
|
| [29] |
Imayoshi I, Sakamoto M, Ohtsuka T, et al. (2008) Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nat Neurosci 11: 1153-1161. doi: 10.1038/nn.2185
|
| [30] |
Mohapel P, Leanza G, Kokaia M, et al. (2005) Forebrain acetylcholine regulates adult hippocampal neurogenesis and learning. Neurobiol Aging 26: 939-946. doi: 10.1016/j.neurobiolaging.2004.07.015
|
| [31] |
Kotani S, Yamauchi T, Teramoto T, et al. (2006) Pharmacological evidence of cholinergic involvement in adult hippocampal neurogenesis in rats. Neuroscience 142: 505-514. doi: 10.1016/j.neuroscience.2006.06.035
|
| [32] |
Mesulam MM, Mufson EJ, Wainer BH, Levey AI (1983) Central cholinergic pathways in the rat: an overview based on an alternative nomenclature (Ch1-Ch6). Neuroscience 10: 1185-1201. doi: 10.1016/0306-4522(83)90108-2
|
| [33] |
Perry E, Walker M, Grace J, et al. (1999) Acetylcholine in mind: a neurotransmitter correlate of consciousness? Trend Neurosci 22: 273-280. doi: 10.1016/S0166-2236(98)01361-7
|
| [34] |
Sarter M, Parikh V (2005) Choline transporters, cholinergic transmission and cognition. Nat Neurosci 6: 48-56. doi: 10.1038/nrn1588
|
| [35] | Mitsushima D, Takahashi T (2011) Contextual learning requires synaptic AMPA receptor delivery in the hippocampus: effect of delivery blocking in behaving rats. Cold Spring Harbor Laboratory Abstr, Synapses: from molecules to circuits & behavior 90. |
| [36] | Taniguchi H, Ishikawa J, Mitsushima D (2014) Real-time change in the firing rate of hippocampal CA1 neurons before, during, and after the exposure to a specific episode. J Physiol Sci 64: S244. |
| [37] | Mizuno J, Mitsushima D (2013) A possible location of contextual memory: CA1 subfield and laterality of learning-dependent synaptic delivery of AMPA receptors. Soc Neurosci Abstr862.12. |
| [38] | Mizuno J, Sakimoto Y, Kida H, et al. (2014) Learning-dependent synaptic plasticity at CA1 synapses: laterality and a possible location of contextual memory in the hippocampus. Soc Neurosci Abstr 754.11. |
| [39] | Sakimoto Y, Mitsushima D (2014) Learning-dependent synaptic diversity in hippocampal CA1 neurons: encoding of context but not retrieval induces rapid plasticity at excitatory and inhibitory synapses. Soc Neurosci Abstr 754.07. |
| [40] | Takase K, Sakimoto Y, Kimura F, et al. (2014) Developmental trajectory of contextual learning and 24-h acetylcholine release in the hippocampus. Sci Rep 4: 3738. |
| [41] |
Coyle JT, Price DL, DeLong MR (1983) Alzheimer's disease: a disorder of cortical cholinergic innervation. Science 219: 1184-1190. doi: 10.1126/science.6338589
|
| [42] |
Ferri CP, Prince M, Brayne C, et al. (2005) Global prevalence of dementia: a Delphi consensus study. Lancet 366: 2112-2117. doi: 10.1016/S0140-6736(05)67889-0
|
| [43] |
Mount C, Downtown D (2006) Alzheimer disease: progress or profit? Nat Med 12:780-784. doi: 10.1038/nm0706-780
|
| [44] |
Cummings JL (2004) Alzheimer's disease. New Engl J Med, 351: 56-67. doi: 10.1056/NEJMra040223
|
| [45] |
McCurry SM, Logsdon RG, Vitiello MV, et al. (2004) Treatment of sleep and nighttime disturbances in Alzheimer's disease: a behavior management approach. Sleep Med 5: 373-377. doi: 10.1016/j.sleep.2003.11.003
|
| [46] |
Starkstein SE, Jorge R, Mizrahi R, et al. (2005) The construct of minor and major depression in Alzheimer's disease. Am J Psychiatry 162: 2086-2093. doi: 10.1176/appi.ajp.162.11.2086
|
| [47] |
Petersen RC, Thomas RG, Grundman M, et al. (2005) Vitamin E and donepezil for the treatment of mild cognitive impairment. New Engl J Med 352: 2379-2388. doi: 10.1056/NEJMoa050151
|
| [48] |
Winblad B, Kilander L, Eriksson S, et al. (2006) Donepezil in patients with severe Alzheimer’s disease: double-blind, parallel-group, placebo-controlled study. Lancet 367: 1057-1065. doi: 10.1016/S0140-6736(06)68350-5
|
| [49] | Mitsushima D, Takase K, Funabashi T, et al. (2008) Gonadal steroid hormones maintain the stress-induced acetylcholine release in the hippocampus: simultaneous measurements of the extracellular acetylcholine and serum corticosterone levels in the same subjects. Endocrinology149: 802-811. |
Figures(5)

Dai Mitsushima. Contextual Learning Requires Functional Diversity at Excitatory and Inhibitory Synapses onto CA1 Pyramidal Neurons[J]. AIMS Neuroscience, 2015, 2(1): 7-17. doi: 10.3934/Neuroscience.2015.1.7







 DownLoad:
DownLoad: