Citation: Yu Murayama. Inequality, mobility, and growth[J]. National Accounting Review, 2019, 1(1): 62-70. doi: 10.3934/NAR.2019.1.62

| [1] | Alesina A, Rodrik D (1994) Distributive Politics and Economic Growth. Q J Econ 109: 465-490. |
| [2] | Andrews D, Leigh A (2009) More Inequality, Less Social Mobility. Appl Econ Lett 16: 1489-1492. |
| [3] | Barro RJ (2000) Inequality and Growth in a Panel of Countries. J Econ Growth 5: 5-32. |
| [4] | Getachew YY (2016) Credit constraints, growth and inequality dynamics. Econ Model 54: 364-376. |
| [5] | Hassler J, Rodríguez Mora JV, Zeira J (2007) Inequality and mobility. J Econ Growth 12: 235-259. |
| [6] | Maoz YD, Moav O (1999) Intergenerational mobility and the process of development. Econ J 109: 677-697. |
| [7] |
Perotti R (1996) Growth, Income Distribution, and Democracy: What the Data Say. J Econ Growth 1: 149-187. doi: 10.1007/BF00138861

|
| [8] | Persson T, Tabellini G (1994) Is Inequality Harmful for Growth? Am Econ Rev 84: 600-621. |
| [9] | Rodriguez JP, Rodriguez JG, Salas R (2008) A study on the relationship between economic inequality and mobility. Econ Lett 99: 111-114. |
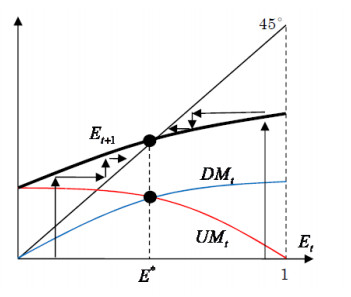
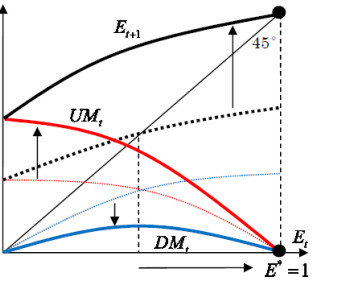
Figures(3)

Yu Murayama. Inequality, mobility, and growth[J]. National Accounting Review, 2019, 1(1): 62-70. doi: 10.3934/NAR.2019.1.62







 DownLoad:
DownLoad: