Citation: Lucie Semenec, Ashley E Franks. Delving through electrogenic biofilms: from anodes to cathodes to microbes[J]. AIMS Bioengineering, 2015, 2(3): 222-248. doi: 10.3934/bioeng.2015.3.222

| [1] |
Bogino PC, Oliva MD, Sorroche FG, et al. (2013) The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations. Int J Mol Sci 14: 15838-15859. doi: 10.3390/ijms140815838

|
| [2] | Aracic S, Semenec L, Franks AE (2014) Investigating microbial activities of electrode-associated microorganisms in real-time. Front Microbiol 5: 663. |
| [3] |
Wang H, Ren ZJ (2013) A comprehensive review of microbial electrochemical systems as a platform technology. Biotechnol Adv 31: 1796-1807. doi: 10.1016/j.biotechadv.2013.10.001
|
| [4] |
Rosenbaum MA, Franks AE (2014) Microbial catalysis in bioelectrochemical technologies: status quo, challenges and perspectives. Appl Microbiol Biotechnol 98: 509-518. doi: 10.1007/s00253-013-5396-6
|
| [5] | Dulon S, Parot S, Delia ML, et al. (2007) Electroactive biofilms: new means for electrochemistry. J Appl Electrochem 37: 173-179. |
| [6] |
Mathis BJ, Marshall CW, Milliken CE, et al. (2008) Electricity generation by thermophilic microorganisms from marine sediment. Appl Microbiol Biol 78: 147-155. doi: 10.1007/s00253-007-1266-4
|
| [7] |
Tender LM, Reimers CE, Stecher HA, et al. (2002) Harnessing microbially generated power on the seafloor. Nat Biotechnol 20: 821-825. doi: 10.1038/nbt716
|
| [8] |
Angenent LT, Karim K, Al-Dahhan MH, et al. (2004) Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol 22: 477-485. doi: 10.1016/j.tibtech.2004.07.001
|
| [9] | Logan BE (2005) Simultaneous wastewater treatment and biological electricity generation. Water Sci Technol 52: 31-37. |
| [10] |
Min B, Kim JR, Oh SE, et al. (2005) Electricity generation from swine wastewater using microbial fuel cells. Water Res 39: 4961-4968. doi: 10.1016/j.watres.2005.09.039
|
| [11] | Flemming HC, Wingender J (2010) The biofilm matrix. Nat Rev Microbiol 8: 623-633. |
| [12] | Borole AP, Reguera G, Ringeisen B, et al. (2012) Electroactive biofilms: Current status and future research needs (vol 4, pg 4813, 2011). Energy Environ Sci 5: 9945-9945. |
| [13] |
Baranitharan E, Khan MR, Prasad DMR, et al. (2015) Effect of biofilm formation on the performance of microbial fuel cell for the treatment of palm oil mill effluent. Bioproc Biosyst Eng 38: 15-24. doi: 10.1007/s00449-014-1239-9
|
| [14] |
Richter H, Nevin KP, Jia HF, et al. (2009) Cyclic voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel cell anodes indicates possible roles of OmcB, OmcZ, type IV pili, and protons in extracellular electron transfer. Energy Environ Sci 2: 506-516. doi: 10.1039/b816647a
|
| [15] |
Shi L, Richardson DJ, Wang Z, et al. (2009) The roles of outer membrane cytochromes of Shewanella and Geobacter in extracellular electron transfer. Environ Microbiol Rep 1: 220-227. doi: 10.1111/j.1758-2229.2009.00035.x
|
| [16] |
Malvankar NS, Lovley DR (2012) Microbial Nanowires: A New Paradigm for Biological Electron Transfer and Bioelectronics. Chemsuschem 5: 1039-1046. doi: 10.1002/cssc.201100733
|
| [17] |
Dheilly A, Linossier I, Darchen A, et al. (2008) Monitoring of microbial adhesion and biofilm growth using electrochemical impedancemetry. Appl Microbiol Biotechnol 79: 157-164. doi: 10.1007/s00253-008-1404-7
|
| [18] |
Malvankar NS, Vargas M, Nevin KP, et al. (2011) Tunable metallic-like conductivity in microbial nanowire networks. Nat Nanotechnol 6: 573-579. doi: 10.1038/nnano.2011.119
|
| [19] |
Malvankar NS, Lau J, Nevin KP, et al. (2012) Electrical Conductivity in a Mixed-Species Biofilm. Appl Environ Microbiol 78: 5967-5971. doi: 10.1128/AEM.01803-12
|
| [20] |
Wagner RC, Call DF, Logan BE (2010) Optimal set anode potentials vary in bioelectrochemical systems. Environ Sci Technol 44: 6036-6041. doi: 10.1021/es101013e
|
| [21] |
Kiely PD, Regan JM, Logan BE (2011) The electric picnic: synergistic requirements for exoelectrogenic microbial communities. Curr Opin Biotechnol 22: 378-385. doi: 10.1016/j.copbio.2011.03.003
|
| [22] |
Nevin KP, Richter H, Covalla SF, et al. (2008) Power output and columbic efficiencies from biofilms of Geobacter sulfurreducens comparable to mixed community microbial fuel cells. Environ Microbiol 10: 2505-2514. doi: 10.1111/j.1462-2920.2008.01675.x
|
| [23] |
Biffinger JC, Pietron J, Ray R, et al. (2007) A biofilm enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and oxygen reduction cathodes. Biosens Bioelectron 22: 1672-1679. doi: 10.1016/j.bios.2006.07.027
|
| [24] |
Lanthier M, Gregory KB, Lovley DR (2008) Growth with high planktonic biomass in Shewanella oneidensis fuel cells. Fems Microbiol Lett 278: 29-35. doi: 10.1111/j.1574-6968.2007.00964.x
|
| [25] | Ringeisen BR, Henderson E, Wu PK, et al. (2006) High Power Density from a Miniature Microbial Fuel Cell Using Shewanella oneidensis DSP10 (vol 40, pg 2629, 2006). Environ Sci Technol 47: 2143-2143. |
| [26] |
Rosenbaum MA, Bar HY, Beg QK, et al. (2011) Shewanella oneidensis in a lactate-fed pure-culture and a glucose-fed co-culture with Lactococcus lactis with an electrode as electron acceptor. Bioresource Technol 102: 2623-2628. doi: 10.1016/j.biortech.2010.10.033
|
| [27] |
Rabaey K, Boon N, Hofte M, et al. (2005) Microbial phenazine production enhances electron transfer in biofuel cells. Environ Sci Technol 39: 3401-3408. doi: 10.1021/es048563o
|
| [28] |
Cordas CM, Guerra LT, Xavier C, et al. (2008) Electroactive biofilms of sulphate reducing bacteria. Electrochimica Acta 54: 29-34. doi: 10.1016/j.electacta.2008.02.041
|
| [29] |
Kang CS, Eaktasang N, Kwon DY, et al. (2014) Enhanced current production by Desulfovibrio desulfuricans biofilm in a mediator-less microbial fuel cell. Bioresource Technol 165: 27-30. doi: 10.1016/j.biortech.2014.03.148
|
| [30] |
Wrighton KC, Thrash JC, Melnyk RA, et al. (2011) Evidence for Direct Electron Transfer by a Gram-Positive Bacterium Isolated from a Microbial Fuel Cell. Appl Environ Microbiol 77: 7633-7639. doi: 10.1128/AEM.05365-11
|
| [31] |
Marshall CW, May HD (2009) Electrochemical evidence of direct electrode reduction by a thermophilic Gram-positive bacterium, Thermincola ferriacetica. Energy Environ Sci 2: 699-705. doi: 10.1039/b823237g
|
| [32] |
Chae KJ, Choi MJ, Lee JW, et al. (2009) Effect of different substrates on the performance, bacterial diversity, and bacterial viability in microbial fuel cells. Bioresource Technol 100: 3518-3525. doi: 10.1016/j.biortech.2009.02.065
|
| [33] |
Jung S, Regan JM (2007) Comparison of anode bacterial communities and performance in microbial fuel cells with different electron donors. Appl Microbiol Biotechnol 77: 393-402. doi: 10.1007/s00253-007-1162-y
|
| [34] |
Holmes DE, Bond DR, O'Neil RA, et al. (2004) Microbial communities associated with electrodes harvesting electricity from a variety of aquatic sediments. Microb Ecol 48: 178-190. doi: 10.1007/s00248-003-0004-4
|
| [35] |
Shrestha PM, Malvankar NS, Werner JJ, et al. (2014) Correlation between microbial community and granule conductivity in anaerobic bioreactors for brewery wastewater treatment. Bioresour Technol 174: 306-310. doi: 10.1016/j.biortech.2014.10.004
|
| [36] |
Yates MD, Kiely PD, Call DF, et al. (2012) Convergent development of anodic bacterial communities in microbial fuel cells. ISME J 6: 2002-2013. doi: 10.1038/ismej.2012.42
|
| [37] |
Werner JJ, Knights D, Garcia ML, et al. (2011) Bacterial community structures are unique and resilient in full-scale bioenergy systems. Proc Natl Acad Sci U S A 108: 4158-4163. doi: 10.1073/pnas.1015676108
|
| [38] |
Franks AE, Nevin KP, Glaven RH, et al. (2010) Microtoming coupled to microarray analysis to evaluate the spatial metabolic status of Geobacter sulfurreducens biofilms. Isme J 4: 509-519. doi: 10.1038/ismej.2009.137
|
| [39] |
Franks AE, Nevin KP, Jia HF, et al. (2009) Novel strategy for three-dimensional real-time imaging of microbial fuel cell communities: monitoring the inhibitory effects of proton accumulation within the anode biofilm. Energy Environ Sci 2: 113-119. doi: 10.1039/B816445B
|
| [40] | Read ST, Dutta P, Bond PL, et al. (2010) Initial development and structure of biofilms on microbial fuel cell anodes. Bmc Microbiol 10. |
| [41] |
Jana PS, Katuri K, Kavanagh P, et al. (2014) Charge transport in films of Geobacter sulfurreducens on graphite electrodes as a function of film thickness. Phys Chem Chem Phys 16: 9039-9046. doi: 10.1039/c4cp01023j
|
| [42] |
Reguera G, Nevin KP, Nicoll JS, et al. (2006) Biofilm and nanowire production leads to increased current in Geobacter sulfurreducens fuel cells. Appl Environ Microbiol 72: 7345-7348. doi: 10.1128/AEM.01444-06
|
| [43] |
Robuschi L, Tomba JP, Schrott GD, et al. (2013) Spectroscopic Slicing to Reveal Internal Redox Gradients in Electricity-Producing Biofilms. Angew Chem Int Edit 52: 925-928. doi: 10.1002/anie.201205440
|
| [44] |
Schrott GD, Ordonez MV, Robuschi L, et al. (2014) Physiological Stratification in Electricity-Producing Biofilms of Geobacter sulfurreducens. Chemsuschem 7: 598-603. doi: 10.1002/cssc.201300605
|
| [45] |
Pfeffer C, Larsen S, Song J, et al. (2012) Filamentous bacteria transport electrons over centimetre distances. Nature 491: 218-221. doi: 10.1038/nature11586
|
| [46] |
Lovley DR (2012) Electromicrobiology. Annu Rev Microbiol 66: 391-409. doi: 10.1146/annurev-micro-092611-150104
|
| [47] |
Semenec L, Franks AE (2014) The microbiology of microbial electrolysis cells. Microbiol Aust 35: 201-206. doi: 10.1071/MA14065
|
| [48] |
Mehta T, Coppi MV, Childers SE, et al. (2005) Outer membrane c-type cytochromes required for Fe(III) and Mn(IV) oxide reduction in Geobacter sulfurreducens. Appl Environ Microbiol 71: 8634-8641. doi: 10.1128/AEM.71.12.8634-8641.2005
|
| [49] |
Reguera G, McCarthy KD, Mehta T, et al. (2005) Extracellular electron transfer via microbial nanowires. Nature 435: 1098-1101. doi: 10.1038/nature03661
|
| [50] |
Coursolle D, Baron DB, Bond DR, et al. (2010) The Mtr Respiratory Pathway Is Essential for Reducing Flavins and Electrodes in Shewanella oneidensis. J Bacteriol 192: 467-474. doi: 10.1128/JB.00925-09
|
| [51] | Kotloski NJ, Gralnick JA (2013) Flavin Electron Shuttles Dominate Extracellular Electron Transfer by Shewanella oneidensis. Mbio 4. pii: e00553-12. |
| [52] |
Malvankar NS, Lovley DR (2014) Microbial nanowires for bioenergy applications. Curr Opin Biotechnol 27: 88-95. doi: 10.1016/j.copbio.2013.12.003
|
| [53] |
Giltner CL, Nguyen Y, Burrows LL (2012) Type IV Pilin Proteins: Versatile Molecular Modules. Microbiol Mol Biol Rev 76: 740-772. doi: 10.1128/MMBR.00035-12
|
| [54] |
Giltner CL, van Schaik EJ, Audette GF, et al. (2006) The Pseudomonas aeruginosa type IV pilin receptor binding domain functions as an adhesin for both biotic and abiotic surfaces. Mol Microbiol 59: 1083-1096. doi: 10.1111/j.1365-2958.2005.05002.x
|
| [55] | Gorby YA, Yanina S, McLean JS, et al. (2009) Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other microorganisms (vol 103, pg 11358, 2006). Proc Nat Acad Sci U S A 106: 9535-9535. |
| [56] |
Malvankar NS, Tuominen MT, Lovley DR (2012) Biofilm conductivity is a decisive variable for high-current-density Geobacter sulfurreducens microbial fuel cells. Energy Environ Sci 5: 5790-5797. doi: 10.1039/c2ee03388g
|
| [57] | El-Naggar MY, Wanger G, Leung KM, et al. (2011) Electrical transport along bacterial nanowires from Shewanella oneidensis MR-1. Proc Natl Acad Sci U S A. 107: 18127-31. |
| [58] |
Gorby YA, Yanina S, McLean JS, et al. (2006) Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other microorganisms. Proc Natl Acad Sci U S A 103: 11358-11363. doi: 10.1073/pnas.0604517103
|
| [59] |
Pirbadian S, El-Naggar MY (2012) Multistep hopping and extracellular charge transfer in microbial redox chains. Phys Chem Chem Phys 14: 13802-13808. doi: 10.1039/c2cp41185g
|
| [60] |
Lovley DR, Malvankar NS (2015) Seeing is believing: novel imaging techniques help clarify microbial nanowire structure and function. Environ Microbiol 17: 2209-15. doi: 10.1111/1462-2920.12708
|
| [61] |
Liu X, Tremblay PL, Malvankar NS, et al. (2014) A Geobacter sulfurreducens Strain Expressing Pseudomonas aeruginosa Type IV Pili Localizes OmcS on Pili but Is Deficient in Fe(III) Oxide Reduction and Current Production. Appl Environ Microbiol 80: 1219-1224. doi: 10.1128/AEM.02938-13
|
| [62] |
Lovley DR (2012) Long-range electron transport to Fe(III) oxide via pili with metallic-like conductivity. Biochem Soc Trans 40: 1186-1190. doi: 10.1042/BST20120131
|
| [63] |
Voordeckers JW, Kim BC, Izallalen M, et al. (2010) Role of Geobacter sulfurreducens outer surface c-type cytochromes in reduction of soil humic acid and anthraquinone-2,6-disulfonate. Appl Environ Microbiol 76: 2371-2375. doi: 10.1128/AEM.02250-09
|
| [64] | Vargas M, Malvankar NS, Tremblay PL, et al. (2013) Aromatic Amino Acids Required for Pili Conductivity and Long-Range Extracellular Electron Transport in Geobacter sulfurreducens. Mbio 4: e00105-13. |
| [65] | Malvankar NS, Vargas M, Nevin K, et al. (2015) Structural basis for metallic-like conductivity in microbial nanowires. Mbio 6: e00084. |
| [66] |
Yan H, Chuang C, Zhugayevych A, et al. (2015) Inter-aromatic distances in Geobacter sulfurreducens pili relevant to biofilm charge transport. Adv Mater 27: 1908-1911. doi: 10.1002/adma.201404167
|
| [67] |
Pirbadian S, Barchinger SE, Leung KM, et al. (2014) Shewanella oneidensis MR-1 nanowires are outer membrane and periplasmic extensions of the extracellular electron transport components. Proc Natl Acad Sci U S A 111: 12883-12888. doi: 10.1073/pnas.1410551111
|
| [68] |
Richter K, Schicklberger M, Gescher J (2012) Dissimilatory Reduction of Extracellular Electron Acceptors in Anaerobic Respiration. Appl Environ Microbiol 78: 913-921. doi: 10.1128/AEM.06803-11
|
| [69] |
Bonanni PS, Schrott GD, Robuschi L, et al. (2012) Charge accumulation and electron transfer kinetics in Geobacter sulfurreducens biofilms. Energy Environ Sci 5: 6188-6195. doi: 10.1039/c2ee02672d
|
| [70] |
Malvankar NS, Tuominen MT, Lovley DR (2012) Lack of cytochrome involvement in long-range electron transport through conductive biofilms and nanowires of Geobacter sulfurreducens. Energy Environ Sci 5: 8651-8659. doi: 10.1039/c2ee22330a
|
| [71] |
Leang C, Qian X, Mester T, et al. (2010) Alignment of the c-type cytochrome OmcS along pili of Geobacter sulfurreducens. Appl Environ Microbiol 76: 4080-4084. doi: 10.1128/AEM.00023-10
|
| [72] |
Nevin KP, Kim BC, Glaven RH, et al. (2009) Anode Biofilm Transcriptomics Reveals Outer Surface Components Essential for High Density Current Production in Geobacter sulfurreducens Fuel Cells. Plos One 4: e5628. doi: 10.1371/journal.pone.0005628
|
| [73] |
Inoue K, Leang C, Franks AE, et al. (2011) Specific localization of the c-type cytochrome OmcZ at the anode surface in current-producing biofilms of Geobacter sulfurreducens. Environ Microbiol Rep 3: 211-217. doi: 10.1111/j.1758-2229.2010.00210.x
|
| [74] |
Rollefson JB, Stephen CS, Tien M, et al. (2011) Identification of an Extracellular Polysaccharide Network Essential for Cytochrome Anchoring and Biofilm Formation in Geobacter sulfurreducens. J Bacteriol 193: 1023-1033. doi: 10.1128/JB.01092-10
|
| [75] |
Pierson LS 3rd, Pierson EA (2010) Metabolism and function of phenazines in bacteria: impacts on the behavior of bacteria in the environment and biotechnological processes. Appl Microbiol Biotechnol 86: 1659-1670. doi: 10.1007/s00253-010-2509-3
|
| [76] |
Dietrich LEP, Okegbe C, Price-Whelan A, et al. (2013) Bacterial Community Morphogenesis Is Intimately Linked to the Intracellular Redox State. J Bacteriol 195: 1371-1380. doi: 10.1128/JB.02273-12
|
| [77] |
Pham TH, Boon N, De Maeyer K, et al. (2008) Use of Pseudomonas species producing phenazine-based metabolites in the anodes of microbial fuel cells to improve electricity generation. Appl Microbiol Biotechnol 80: 985-993. doi: 10.1007/s00253-008-1619-7
|
| [78] |
Shen HB, Yong XY, Chen YL, et al. (2014) Enhanced bioelectricity generation by improving pyocyanin production and membrane permeability through sophorolipid addition in Pseudomonas aeruginosa-inoculated microbial fuel cells. Bioresource Technol 167: 490-494. doi: 10.1016/j.biortech.2014.05.093
|
| [79] |
Marsili E, Baron DB, Shikhare ID, et al. (2008) Shewanella Secretes flavins that mediate extracellular electron transfer. Proc Natl Acad Sci U S A 105: 3968-3973. doi: 10.1073/pnas.0710525105
|
| [80] |
Bond DR, Lovley DR (2003) Electricity production by Geobacter sulfurreducens attached to electrodes. Appl Environ Microbiol 69: 1548-1555. doi: 10.1128/AEM.69.3.1548-1555.2003
|
| [81] |
Okamoto A, Nakamura R, Nealson KH, et al. (2014) Bound Flavin Model Suggests Similar Electron-Transfer Mechanisms in Shewanella and Geobacter. Chemelectrochem 1: 1808-1812. doi: 10.1002/celc.201402151
|
| [82] |
Wei JC, Liang P, Cao XX, et al. (2010) A New Insight into Potential Regulation on Growth and Power Generation of Geobacter sulfurreducens in Microbial Fuel Cells Based on Energy Viewpoint. Environ Sci Technol 44: 3187-3191. doi: 10.1021/es903758m
|
| [83] |
Parot S, Delia ML, Bergel A (2008) Forming electrochemically active biofilms from garden compost under chronoamperometry. Bioresource Technol 99: 4809-4816. doi: 10.1016/j.biortech.2007.09.047
|
| [84] |
Torres CI, Krajmalnik-Brown R, Parameswaran P, et al. (2009) Selecting Anode-Respiring Bacteria Based on Anode Potential: Phylogenetic, Electrochemical, and Microscopic Characterization. Environ Sci Technol 43: 9519-9524. doi: 10.1021/es902165y
|
| [85] |
Zhu XP, Yates MD, Hatzell MC, et al. (2014) Microbial Community Composition Is Unaffected by Anode Potential. Environ Sci Technol 48: 1352-1358. doi: 10.1021/es404690q
|
| [86] |
Ishii S, Suzuki S, Norden-Krichmar TM, et al. (2014) Microbial population and functional dynamics associated with surface potential and carbon metabolism. Isme J 8: 963-978. doi: 10.1038/ismej.2013.217
|
| [87] | Levar CE, Chan CH, Mehta-Kolte MG, et al. (2014) An Inner Membrane Cytochrome Required Only for Reduction of High Redox Potential Extracellular Electron Acceptors. Mbio 5. |
| [88] |
Zhu XP, Yates MD, Logan BE (2012) Set potential regulation reveals additional oxidation peaks of Geobacter sulfurreducens anodic biofilms. Electrochem Comm 22: 116-119. doi: 10.1016/j.elecom.2012.06.013
|
| [89] |
Kumar A, Siggins A, Katuri K, et al. (2013) Catalytic response of microbial biofilms grown under fixed anode potentials depends on electrochemical cell configuration. Chem Eng J 230: 532-536. doi: 10.1016/j.cej.2013.06.044
|
| [90] |
Patil SA, Harnisch F, Koch C, et al. (2011) Electroactive mixed culture derived biofilms in microbial bioelectrochemical systems: The role of pH on biofilm formation, performance and composition. Bioresource Technol 102: 9683-9690. doi: 10.1016/j.biortech.2011.07.087
|
| [91] |
Fu Q, Kobayashi H, Kuramochi Y, et al. (2013) Bioelectrochemical analyses of a thermophilic biocathode catalyzing sustainable hydrogen production. Int J Hydrogen Energ 38: 15638-15645. doi: 10.1016/j.ijhydene.2013.04.116
|
| [92] |
Jafary T, Daud WRW, Ghasemi M, et al. (2015) Biocathode in microbial electrolysis cell; present status and future prospects. Renew Sust Energ Rev 47: 23-33. doi: 10.1016/j.rser.2015.03.003
|
| [93] |
Hou Y, Luo H, Liu G, et al. (2014) Improved hydrogen production in the microbial electrolysis cell by inhibiting methanogenesis using ultraviolet irradiation. Environ Sci Technol 48: 10482-10488. doi: 10.1021/es501202e
|
| [94] |
Logan BE, Call D, Cheng S, et al. (2008) Microbial electrolysis cells for high yield hydrogen gas production from organic matter. Environ Sci Technol 42: 8630-8640. doi: 10.1021/es801553z
|
| [95] |
Babauta JT, Hsu L, Atci E, et al. (2014) Multiple cathodic reaction mechanisms in seawater cathodic biofilms operating in sediment microbial fuel cells. ChemSusChem 7: 2898-2906. doi: 10.1002/cssc.201402377
|
| [96] |
Lovley DR, Nevin KP (2013) Electrobiocommodities: powering microbial production of fuels and commodity chemicals from carbon dioxide with electricity. Curr Opin Biotechnol 24: 385-390. doi: 10.1016/j.copbio.2013.02.012
|
| [97] |
Nevin KP, Hensley SA, Franks AE, et al. (2011) Electrosynthesis of organic compounds from carbon dioxide is catalyzed by a diversity of acetogenic microorganisms. Appl Environ Microbiol 77: 2882-2886. doi: 10.1128/AEM.02642-10
|
| [98] |
Villano M, Aulenta F, Ciucci C, et al. (2010) Bioelectrochemical reduction of CO2 to CH4 via direct and indirect extracellular electron transfer by a hydrogenophilic methanogenic culture. Bioresource Technol 101: 3085-3090. doi: 10.1016/j.biortech.2009.12.077
|
| [99] |
Gregory KB, Bond DR, Lovley DR (2004) Graphite electrodes as electron donors for anaerobic respiration. Environ Microbiol 6: 596-604. doi: 10.1111/j.1462-2920.2004.00593.x
|
| [100] |
Pous N, Puig S, Coma M, et al. (2013) Bioremediation of nitrate-polluted groundwater in a microbial fuel cell. J Chem Technol Biotechnol 88: 1690-1696. doi: 10.1002/jctb.4020
|
| [101] |
Gregory KB, Lovley DR (2005) Remediation and recovery of uranium from contaminated subsurface environments with electrodes. Environ Sci Technol 39: 8943-8947. doi: 10.1021/es050457e
|
| [102] |
Aulenta F, Reale P, Canosa A, et al. (2010) Characterization of an electro-active biocathode capable of dechlorinating trichloroethene and cis-dichloroethene to ethene. Biosens Bioelectron 25: 1796-1802. doi: 10.1016/j.bios.2009.12.033
|
| [103] |
Huang L, Shi Y, Wang N, et al. (2014) Anaerobic/aerobic conditions and biostimulation for enhanced chlorophenols degradation in biocathode microbial fuel cells. Biodegradation 25: 615-632. doi: 10.1007/s10532-014-9686-1
|
| [104] |
Strycharz SM, Woodard TL, Johnson JP, et al. (2008) Graphite electrode as a sole electron donor for reductive dechlorination of tetrachlorethene by Geobacter lovleyi. Appl Environ Microb 74: 5943-5947. doi: 10.1128/AEM.00961-08
|
| [105] |
Thrash JC, Van Trump JI, Weber KA, et al. (2007) Electrochemical stimulation of microbial perchlorate reduction. Environ Sci Technol 41: 1740-1746. doi: 10.1021/es062772m
|
| [106] |
Huang L, Chen J, Quan X, et al. (2010) Enhancement of hexavalent chromium reduction and electricity production from a biocathode microbial fuel cell. Bioprocess Biosyst Eng 33: 937-945. doi: 10.1007/s00449-010-0417-7
|
| [107] |
Tandukar M, Huber SJ, Onodera T, et al. (2009) Biological Chromium(VI) Reduction in the Cathode of a Microbial Fuel Cell. Environ Sci Technol 43: 8159-8165. doi: 10.1021/es9014184
|
| [108] |
He Z, Angenent LT (2006) Application of bacterial biocathodes in microbial fuel cells. Electroanalysis 18: 2009-2015. doi: 10.1002/elan.200603628
|
| [109] | Gregory KB, Bond DR, Lovley DR (2004) Graphite electrodes as electron donors for anaerobic respiration. Environ Microbiol 6: 596-604. |
| [110] |
Gregory KB, Lovley DR (2005) Remediation and recovery of uranium from contaminated subsurface environments with electrodes. Environ Sci Technol 39: 8943-8947. doi: 10.1021/es050457e
|
| [111] |
Zhang W, Zhang Y, Su W, et al. (2014) Effects of cathode potentials and nitrate concentrations on dissimilatory nitrate reductions by Pseudomonas alcaliphila in bioelectrochemical systems. J Environ Sci (China) 26: 885-891. doi: 10.1016/S1001-0742(13)60460-X
|
| [112] |
Cheng S, Xing D, Call DF, et al. (2009) Direct biological conversion of electrical current into methane by electromethanogenesis. Environ Sci Technol 43: 3953-3958. doi: 10.1021/es803531g
|
| [113] |
Aulenta F, Catapano L, Snip L, et al. (2012) Linking Bacterial Metabolism to Graphite Cathodes: Electrochemical Insights into the H2-Producing Capability of Desulfovibrio sp. Chemsuschem 5: 1080-1085. doi: 10.1002/cssc.201100720
|
| [114] |
Rosenbaum M, Aulenta F, Villano M, et al. (2011) Cathodes as electron donors for microbial metabolism: Which extracellular electron transfer mechanisms are involved? Bioresource Technol 102: 324-333. doi: 10.1016/j.biortech.2010.07.008
|
| [115] | Mateju V, Cizinska S, Krejci J, et al. (1992) Biological Water Denitrification—a Review. Enzyme Microb Tech 14: 170-183. |
| [116] |
Park HI, Kim DK, Choi YJ, et al. (2005) Nitrate reduction using an electrode as direct electron donor in a biofilm-electrode reactor. Proc Biochem 40: 3383-3388. doi: 10.1016/j.procbio.2005.03.017
|
| [117] |
Vilar-Sanz A, Puig S, Garcia-Lledo A, et al. (2013) Denitrifying Bacterial Communities Affect Current Production and Nitrous Oxide Accumulation in a Microbial Fuel Cell. Plos One 8: e63460. doi: 10.1371/journal.pone.0063460
|
| [118] |
Wrighton KC, Virdis B, Clauwaert P, et al. (2010) Bacterial community structure corresponds to performance during cathodic nitrate reduction. ISME J 4: 1443-1455. doi: 10.1038/ismej.2010.66
|
| [119] |
Kato S, Hashimoto K, Watanabe K (2012) Microbial interspecies electron transfer via electric currents through conductive minerals. Proc Natl Acad Sci U S A 109: 10042-10046. doi: 10.1073/pnas.1117592109
|
| [120] |
Jourdin L, Freguia S, Donose BC, et al. (2015) Autotrophic hydrogen-producing biofilm growth sustained by a cathode as the sole electron and energy source. Bioelectrochemistry 102: 56-63. doi: 10.1016/j.bioelechem.2014.12.001
|
| [121] |
Croese E, Jeremiasse AW, Marshall IPG, et al. (2014) Influence of setup and carbon source on the bacterial community of biocathodes in microbial electrolysis cells. Enzyme Microb Tech 61-62: 67-75. doi: 10.1016/j.enzmictec.2014.04.019
|
| [122] | Morita M, Malvankar NS, Franks AE, et al. (2011) Potential for Direct Interspecies Electron Transfer in Methanogenic Wastewater Digester Aggregates. Mbio 2. |
| [123] |
Kiely PD, Rader G, Regan JM, et al. (2011) Long-term cathode performance and the microbial communities that develop in microbial fuel cells fed different fermentation endproducts. Bioresour Technol 102: 361-366. doi: 10.1016/j.biortech.2010.05.017
|
| [124] |
Strycharz-Glaven SM, Glaven RH, Wang Z, et al. (2013) Electrochemical investigation of a microbial solar cell reveals a nonphotosynthetic biocathode catalyst. Appl Environ Microbiol 79: 3933-3942. doi: 10.1128/AEM.00431-13
|
| [125] |
Yuan Y, Zhou S, Tang J (2013) In situ investigation of cathode and local biofilm microenvironments reveals important roles of OH- and oxygen transport in microbial fuel cells. Environ Sci Technol 47: 4911-4917. doi: 10.1021/es400045s
|
| [126] | Rozendal RA, Jeremiasse AW, Hamelers HV, et al. (2008) Hydrogen production with a microbial biocathode. Environ Sci Technol 42: 629-634. |
| [127] |
Zaybak Z, Pisciotta JM, Tokash JC, et al. (2013) Enhanced start-up of anaerobic facultatively autotrophic biocathodes in bioelectrochemical systems. J Biotechnol 168: 478-485. doi: 10.1016/j.jbiotec.2013.10.001
|
| [128] |
Leang C, Malvankar NS, Franks AE, et al. (2013) Engineering Geobacter sulfurreducens to produce a highly cohesive conductive matrix with enhanced capacity for current production. Energ Environ Sci 6: 1901-1908. doi: 10.1039/c3ee40441b
|
| [129] |
Huang LP, Regan JM, Quan X (2011) Electron transfer mechanisms, new applications, and performance of biocathode microbial fuel cells. Bioresource Technol 102: 316-323. doi: 10.1016/j.biortech.2010.06.096
|
| [130] |
Strycharz SM, Glaven RH, Coppi MV, et al. (2011) Gene expression and deletion analysis of mechanisms for electron transfer from electrodes to Geobacter sulfurreducens. Bioelectrochemistry 80: 142-150. doi: 10.1016/j.bioelechem.2010.07.005
|
| [131] |
Dantas JM, Campelo LM, Duke NE, et al. (2015) The structure of PccH from Geobacter sulfurreducens - a novel low reduction potential monoheme cytochrome essential for accepting electrons from an electrode. FEBS J 282: 2215-2231. doi: 10.1111/febs.13269
|
| [132] | Milner E, Scott K, Head I, et al. (2014) Electrochemical Investigation of Aerobic Biocathodes at Different Poised Potentials: Evidence for Mediated Extracellular Electron Transfer. 10th Esee: European Symposium on Electrochemical Engineering. Sardinia, Italy. 41: 355-360. |
| [133] |
Dinh HT, Kuever J, Mussmann M, et al. (2004) Iron corrosion by novel anaerobic microorganisms. Nature 427: 829-832. doi: 10.1038/nature02321
|
| [134] |
Lohner ST, Deutzmann JS, Logan BE, et al. (2014) Hydrogenase-independent uptake and metabolism of electrons by the archaeon Methanococcus maripaludis. Isme J 8: 1673-1681. doi: 10.1038/ismej.2014.82
|
| [135] |
Bourdakos N, Marsili E, Mahadevan R (2014) A Defined Co-Culture of Geobacter Sulfurreducens and Escherichia Coli in a Membrane-Less Microbial Fuel Cell. Biotechnol Bioeng 111: 709-718. doi: 10.1002/bit.25137
|
| [136] | Smith JA, Nevin KP, Lovley DR (2015) Syntrophic growth via quinone-mediated interspecies electron transfer. Front Microbiol 6: 121. |
| [137] |
Summers ZM, Fogarty HE, Leang C, et al. (2010) Direct Exchange of Electrons Within Aggregates of an Evolved Syntrophic Coculture of Anaerobic Bacteria. Science 330: 1413-1415. doi: 10.1126/science.1196526
|
| [138] |
Rotaru AE, Shrestha PM, Liu FH, et al. (2014) A new model for electron flow during anaerobic digestion: direct interspecies electron transfer to Methanosaeta for the reduction of carbon dioxide to methane. Energ Environ Sci 7: 408-415. doi: 10.1039/C3EE42189A
|
| [139] |
Parameswaran P, Torres CI, Lee HS, et al. (2009) Syntrophic Interactions Among Anode Respiring Bacteria (ARB) and Non-ARB in a Biofilm Anode: Electron Balances. Biotechnol Bioeng 103: 513-523. doi: 10.1002/bit.22267
|
| [140] |
Stratford JP, Beecroft NJ, Slade RCT, et al. (2014) Anodic microbial community diversity as a predictor of the power output of microbial fuel cells. Bioresource Technol 156: 84-91. doi: 10.1016/j.biortech.2014.01.041
|
| [141] |
Smith JA, Tremblay PL, Shrestha PM, et al. (2014) Going Wireless: Fe(III) Oxide Reduction without Pili by Geobacter sulfurreducens Strain JS-1. Appl Environ Microbiol 80: 4331-4340. doi: 10.1128/AEM.01122-14
|
| [142] | Commault AS, Lear G, Weld RJ (2015) Maintenance of Geobacter-dominated biofilms in microbial fuel cells treating synthetic wastewater. Bioelectrochemistry pii: S1567-5394. |
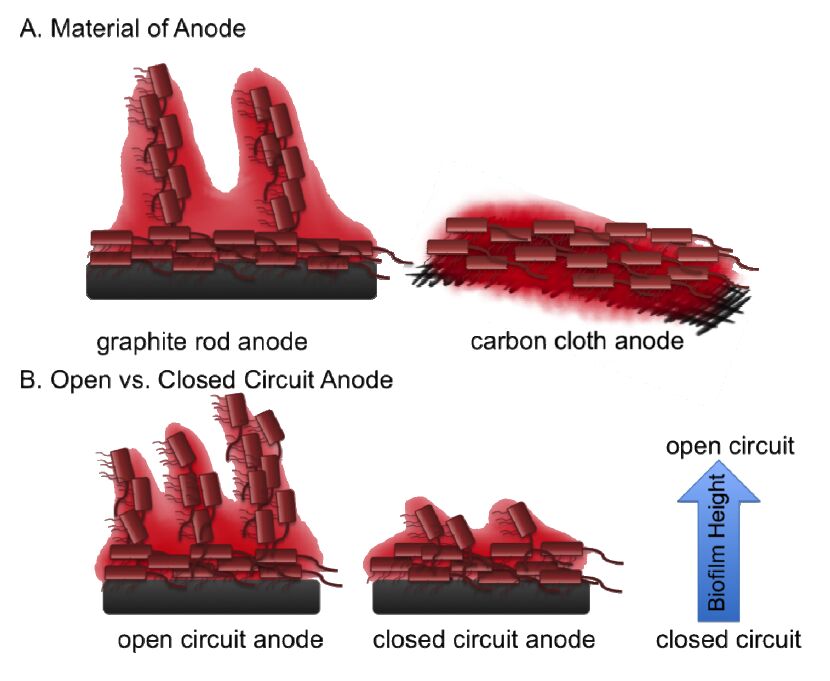
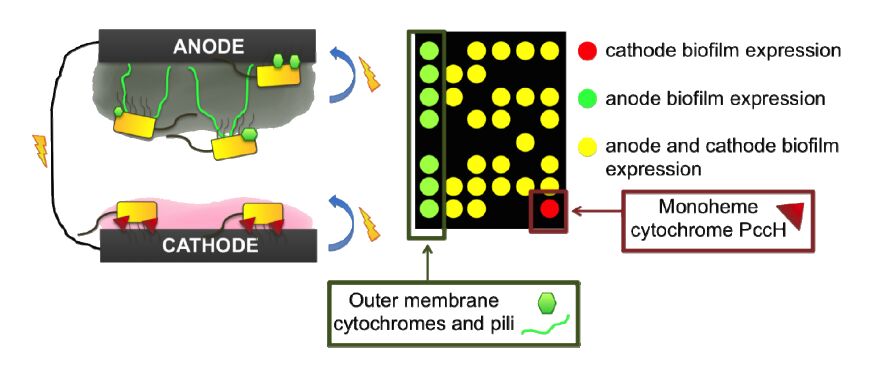
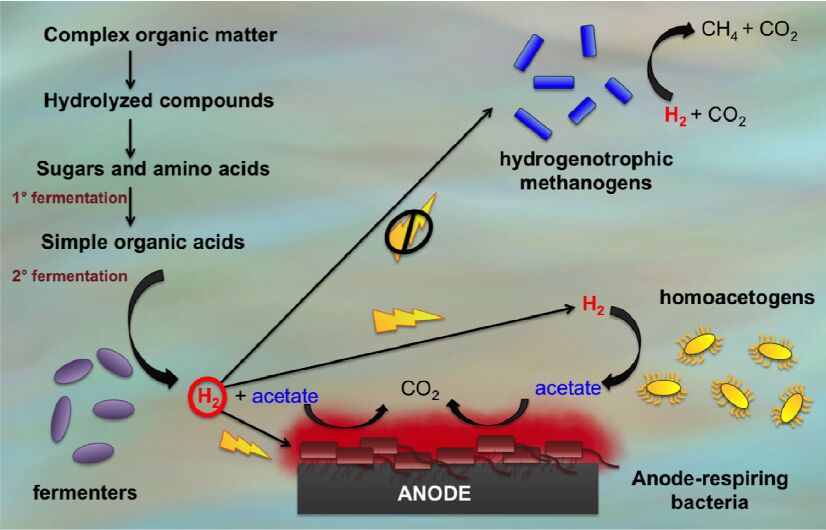
Figures(3)

Lucie Semenec, Ashley E Franks. Delving through electrogenic biofilms: from anodes to cathodes to microbes[J]. AIMS Bioengineering, 2015, 2(3): 222-248. doi: 10.3934/bioeng.2015.3.222







 DownLoad:
DownLoad: