Citation: Kevin R Caffrey, Mari S Chinn, Matthew W Veal. Biomass supply chain management in North Carolina (part 1): predictive model for cropland conversion to biomass feedstocks[J]. AIMS Energy, 2016, 4(2): 256-279. doi: 10.3934/energy.2016.2.256

| [1] | U.S. House (2007) Energy Independence and Security Act of 2007. 100th Congress, 1st session, HR 6.4. |
| [2] | U.S. DOE (2012) Monthly Energy Review: October 2012. US Department of Energy, Energy Information Administration, Independent Statistics & Analysis, DOE/EIA-0035(2012/10). |
| [3] | McKendry P (2002) Energy production from biomass (part 2): conversion technologies. Bioresource Technology 83.1: 47-54. |
| [4] | U.S. DOE (2005) Biomass as Feedstock for a Bioenergy and Bioproducts Industry: The Technical Feasibility of a Billion-Ton Annual Supply. R.D. Perlack, B.J. Stokes, and D.C. Erbach (Leads). United States Department of Energy, Oak Ridge National Laboratory, ORNL/TM-2005/66. |
| [5] | U.S. DOE (2011) U.S. Billion-Ton Update: Biomass Supply for a Bioenergy and Bioproducts Industry. R.K. Perlack and B.J. Stokes (Leads), United States Department of Energy, Oak Ridge National Laboratory, ORNL/TM-2011/224. |
| [6] | Green JD, Witt WW, Margin JR (2006) Weed Management in Grass Pastures, Hayfields, and Other Farmstead Sites (AGR-172). University of Kentucky Cooperative Extension Service. |
| [7] | Wright L, Turhollow A (2010) Switchgrass Selection as a “Model” Bioenergy Crop: A History of the Process. Biomass & Bioenergy 34: 851-868. |
| [8] | Greef JM, Deuter M, Jung C, et al. (1997) Genetic diversity of European Miscanthus species revealed by Aflp finger printing. Genetic Resource & Crop Evolution 44: 185-195. |
| [9] | West NM, Matlaga DP, Davis AS (2014) Managing Spread from Rhizome Fragments is Key to Reducing Invasiveness of Miscanthus x giganteus. Invasive Plant Science & Management 7: 517-525. |
| [10] |
Boose AB, Holt JS (1999) Environmental effects on asexual reproduction in Arundo onax. Weed Research 39: 117-127. doi: 10.1046/j.1365-3180.1999.00129.x

|
| [11] | Palmer IE, Gehl RJ, Ranney TG, et al. (2014) Biomass Yield, Nitrogen Response, and Nutrient Uptake of Perennial Bioenergy Grasses in North Carolina. Biomass & Bioenergy 63: 218-228. |
| [12] | Gill JR, Burks PS, Staggenborg SA, et al. (2014) Yield Results and Stability Analysis form the Sorghum Regional Biomass Feedstock Trial. Bioenergy Research 7: 1026-1034. |
| [13] | Whitfield M, Chinn M, Veal M (2012) Processing of materials derived from sweet sorghum for biobased products. Industrial Crops & Products 37: 362-375. |
| [14] | Köppen S, Reinhardt G, Gärtner S (2009) Assessment of energy and greenhouse gas inventories of Sweet Sorghum for first and second generation bioethanol. Environment and Natural Resources Management Working Paper, 30. Food and Agricultural Organization of the United Nations. |
| [15] |
Griffey C, Brooks W, Kurantz M, et al. (2010) Grain composition of Virginia winter barley and implications for use in feed, food, and biofuels production. Journal of Cereal Sciences 51: 41-49. doi: 10.1016/j.jcs.2009.09.004
|
| [16] | Han M, Kang KE, Kim Y, et al. (2013) High efficiency bioethanol production from barley straw using a continuous pretreatment reactor. Process Biochemistry 48: 488-495. |
| [17] | Zentková I, Cvengrošová E (2013) The utilization of rapeseed for biofuels production in the EU. Visegrad Journal on Bioeconomy & Sustainable Development 2: 11-14. |
| [18] | U.S. DOE (2002) Lignocellulosic Biomass to Ethanol Process Design and Economics Utilizing Co-Current Dilute Acid Prehydrolysis and Enzymatic Hydrolysis for Corn Stover. A. Aden, M. Ruth, K. Ibsen, J. Jechura, K. Neeves, J. Sheehan, B. Wallace, L. Montague, A. Slayton, J. Lukas. United States Department of Energy, National Renewable Energy Laboratory, NREL/TP-510-32438. |
| [19] | Searchinger T, Heinlich R, Houghton RA, et al. (2008) Use of U.S. Croplands for Biofuels Increases Greenhouse Gases Through Emissions from Land-Use Change. Science 319: 1238-1240. |
| [20] | Dale VH, Kline KL, Wiens J, et al. (2010) Biofuels: Implications for Land Use and Biodiversity. Ecological Society of America, Biofuels and Sustainability Reports. |
| [21] | Mayer ML (2012) Assessment of Bioenergy Crop Production Feasibility along North Carolina Department of Transportation Highway Right-of-Ways. NCSU MS Thesis. |
| [22] | Babcock BA, Iqbal Z (2014) Using Recent Land Use Change to Validate Land Use Change Models. Center for Agricultural and Rural Development, Iowa State University. Staff Report 14-SR 109. |
| [23] | Deininger K, Byerlee D, Lindsay J, et al. (2011) Rising Global Interest in Farmland: Can it Yield Sustainable and Equitable Benefits? The World Bank, Agriculture and Rural Development. |
| [24] | Crouse D (2003) Realistic yields and nitrogen application factors for North Carolina crops. North Carolina State University, North Carolina Department of Agriculture and Consumer Services, North Carolina Department of Environment and Natural Resources, Natural Resources Conservation Service. Raleigh NC. Available from: http://nutrients.soil.ncsu.edu/yields/. |
| [25] | USDA (2014) Web Soil Survey. Soil Survey Staff, National Resources Conservation Service, United States Department of Agriculture. Available from: http://websoilsurvey.nrcs.usda.gov. Accessed: October 2014. |
| [26] | USDA (2014) Cropland Data Layer 2013. Published crop-specific data layer [online]. United States Department of Agriculture, National Agricultural Statistics Service. Available from: http://nassgeodata.gmu.edu/CropScape. Accessed: July 2014. |
| [27] |
USDA (2013) 2013 State Agriculture Overview: North Carolina. United States Department of Agriculture, National Agricultural Statistics Service. Available from: http://www.nass.usda.gov/Quick_Stats/Ag_Overview/. Accessed: Nov 2014. |
| [28] | George N, Tungate K, Beec, C, et al. (2010) An Evaluation of winter canola in North Carolina. North Carolina State University, Department of Crop Science, Official Variety Trials. Available from: http://www.ncovt.com/. Accessed: July 2014. |
| [29] | Bullen G, Weddington E (2012) Enterprise Budgets: Corn- Conventional Till-NC, Coastal Plain 2012. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [30] | Bullen G, Weddington E (2012) Enterprise Budgets: Corn- No Till-NC, Coastal Plain 2012. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [31] | Bullen G (2013) Enterprise Budgets: Cotton- Conventional Tillage- 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [32] | Bullen G (2013) Enterprise Budgets: Cotton- Strip Tillage- 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [33] | Bullen G, Jordan D (2013) Enterprise Budgets: Peanuts- Conventional Virginia 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [34] | Bullen G, Jordan D (2013) Enterprise Budgets: Peanuts- Strip Till 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [35] | Bullen G, Dunphy J (2012) Enterprise Budgets: Soybeans- Full Season, Conventional Tillage 2012. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [36] | Bullen G, Dunphy J (2012) Enterprise Budgets: Soybeans- Full Season, No Till 2012. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [37] | Bullen G, Fisher L (2013) Enterprise Budgets: Flue-Cured Tobacco-Hand Harvested Piedmont- 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [38] | Bullen G, Fisher L (2013) Enterprise Budgets: Flue-Cured Tobacco-Machine Harvested Piedmont- 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [39] | Bullen G, Fisher L (2013) Enterprise Budgets: Flue-Cured Tobacco-Machine Harvested East- 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [40] | Bullen G, Weddington E (2012) Enterprise Budgets: Wheat for Grain Conventional 2012. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [41] | Bullen G, Weddington E (2012) Enterprise Budgets: Wheat for Grain No-Till 2012. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [42] |
Bullen G, Little B (2013) Enterprise Budgets: Sweet Potato 2013. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [43] | NCSU (2014) Enterprise Budget: No-Till Grain Sorghum for the Coastal Plain Region of NC. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [44] |
Green JT, Benson GA (2013) Bermuda Grass for Pasture. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [45] |
Green JT, Benson GA (2013) Bermuda Grass for Hay. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [46] |
Green JT, Benson GA (2013) Cool Season Perennial Grass for Pasture. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [47] |
Green JT, Benson GA (2013) Hay Harvest Cost, Large Round Baler. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [48] | U.S. DOE (2009) Uniform-Format Solid Feedstock Supply System: A Commodity-Scale Design to Produce an Infrastructure-Compatible Bulk Solid from Lignocellulosic Biomass. Hess, JR, Wright, CT, Kenney, KL, Searcy, EM. US Department of Energy, Idaho National Laboratory, INL/EXT-09-15423. |
| [49] | Saxe C (2004) Big Bale Storage Losses. University of Wisconsin Extension, November 2004. |
| [50] |
Buchholz T, Volk TA (2011) Improving the Profitability of Willow Crops- Identifying Opportunities with a Crop Budget Model. Bioenergy Research 4: 85-95. doi: 10.1007/s12155-010-9103-5
|
| [51] | Atkinson AD, Rich BA, Tungate KD, et al. (2006) North Carolina Canola Production. North Carolina State University, North Carolina Solar Center & College of Agriculture and Life Sciences. SJS/KEL-9/06-W07 |
| [52] | MB (2012) 2012 Sorghum Program- Murphy-Brown’s Next Step in Broad Strategy to Improve MB Grain Buying. Murphy-Brown Grain, Available from: http://mbgrain.com/ |
| [53] | CC (2014) Current Canola Oil, Meal, and Seed Prices. Canola Council of Canada. Available from: http://www.canolacouncil.org/. |
| [54] | Lee D, Owens VN, Boe A, et al. (2007) Composition of Herbaceous Biomass Feedstocks. SunGrant Initiative, North Central Center, South Dakota State University, SGINC1-07. |
| [55] |
Liebig MA, Schmer MR, Vogel KP, et al. (2008) Soil Carbon Storage by Switchgrass Grown for Bioenergy. Bioenergy Research 1: 215-222. doi: 10.1007/s12155-008-9019-5
|
| [56] | Humbird D, Davis R, Tao L, et al. (2011) Process Design and Economics for Biochemical Conversion of Lignocellulosic Biomass to Ethanol: Dilute-Acid Pretreatment and Enzymatic Hydrolysis of Corn Stover. US Department of Energy, National Renewable Energy Laboratory, NRL/TP-5100-47764. |
| [57] | Glassner D, Hettenhaus J, Schechinger T (1998) Corn stover collection project- a pilot for establishing infrastructure for agricultural residue and other crop collection for biomass processing to ethanol. In: Proc. Bioenergy 1998 Conference, 4-8 October, 1998, Madison, WI, pp. 1100-1110. |
| [58] | Lazarus WF (2009) Machinery Cost Estimates. University of Minnesota Extension. St Paul, MN. Shelf Location: 378.776 A4744 6696 2009-6. |
| [59] | Lazarus WF (2014) Machinery Cost Estimates. University of Minnesota Extension. St. Paul, MN. Available at: https://drive.google.com/a/umn.edu/file/d/0B3psjoooP5QxWWd3a2cwblJCTjQ/view |
| [60] | USDA (2015) Cash Rents by County. United States Department of Agriculture, National Agricultural Statistics Service. August 2014. Available at: www.nass.usda.gov |
| [61] | U.S. DOE (2011) Biomass multi-year program plan. United States Department of Energy, Energy Efficiency & Renewable Energy, Office of Biomass Program. |
| [62] | NCSU (2008) Enterprise Budgets: Hay Production, Harvest and Storage Costs, Round Bales. North Carolina State University, Department of Agricultural and Resource Economics. Available from: http://ag-econ.ncsu.edu/extension/budgets. |
| [63] | U.S. DOE (2014) Southeast Biomass Atlas. United States Department of Energy, SunGrant Initiative Southeastern Regional Center, Oak Ridge National Laboratory, University of Tennessee Center for Renewable Carbon. Available from: http://www.biomassatlas.org/ |
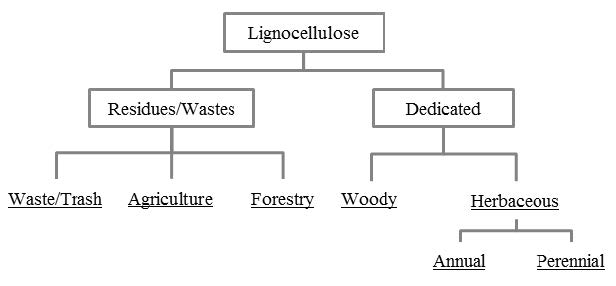
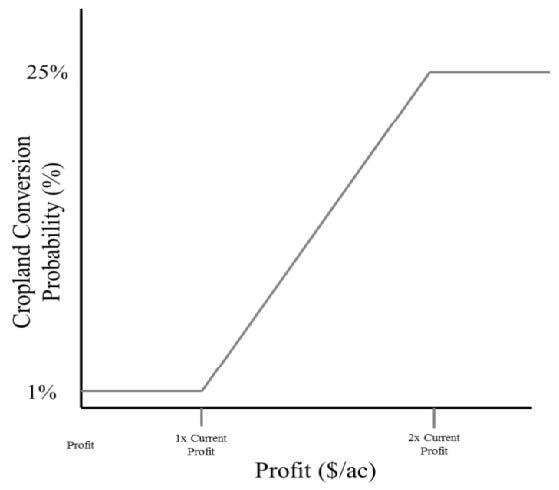

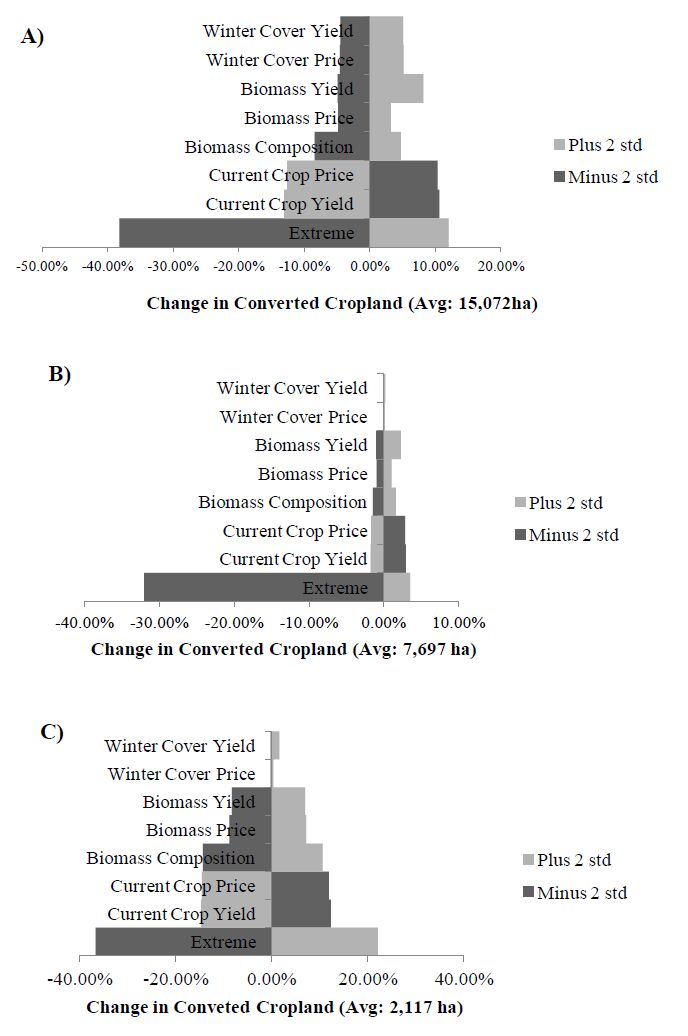
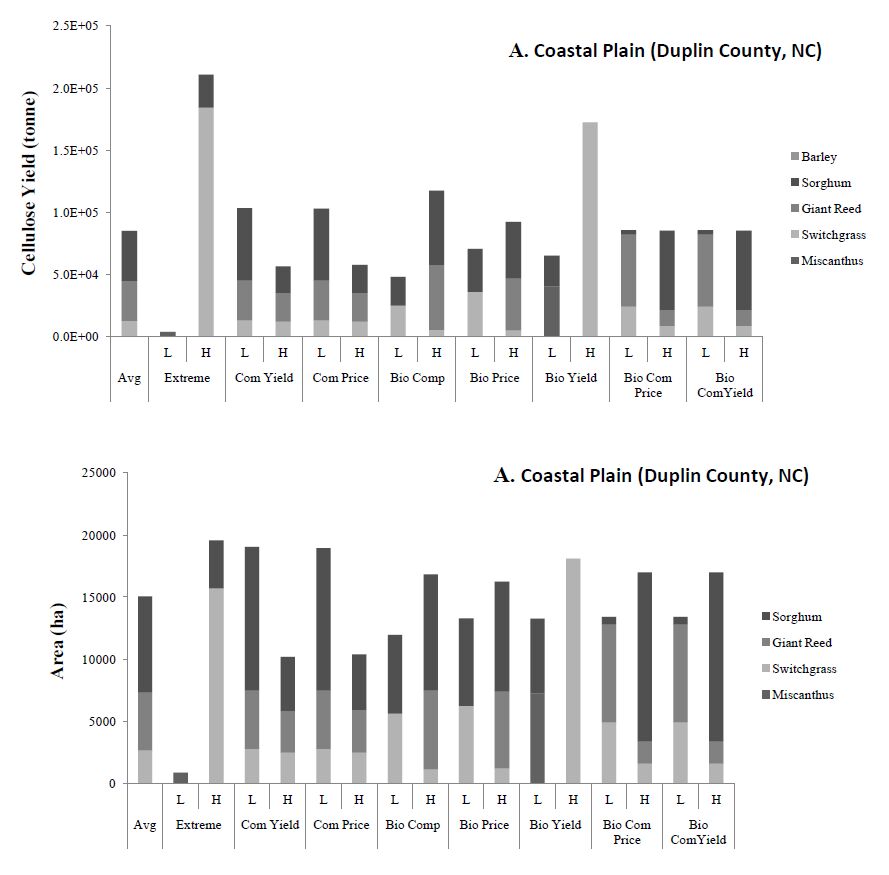
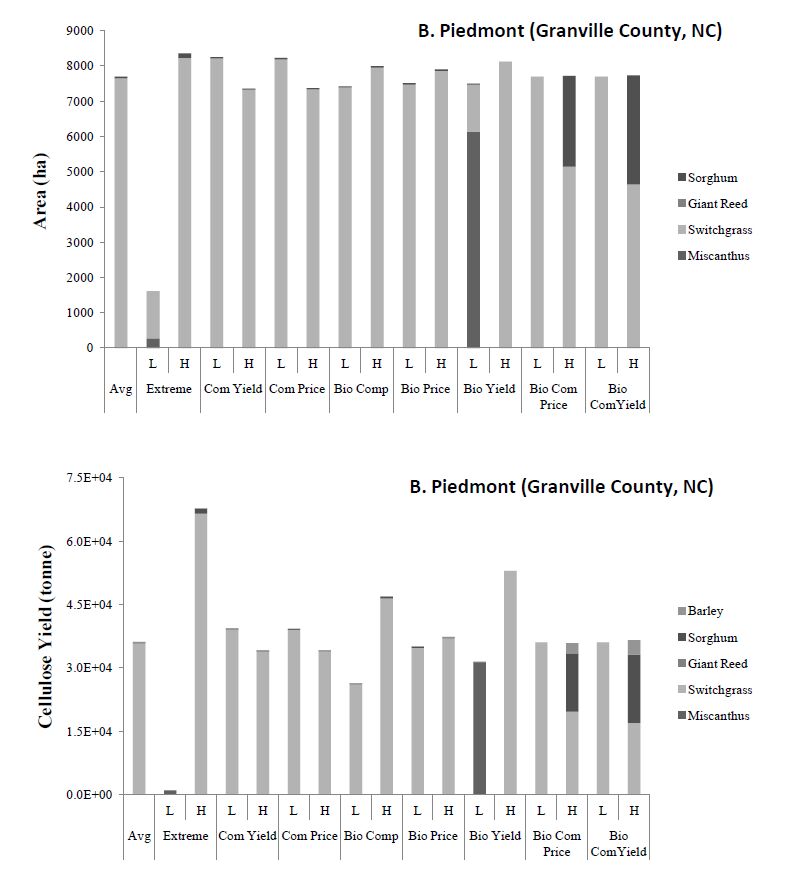
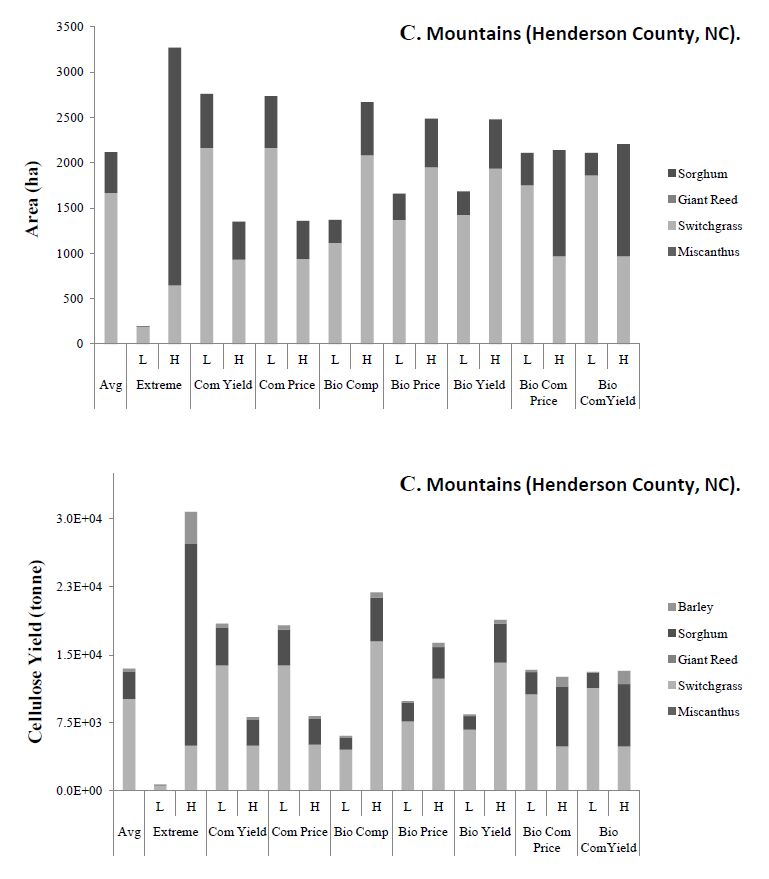
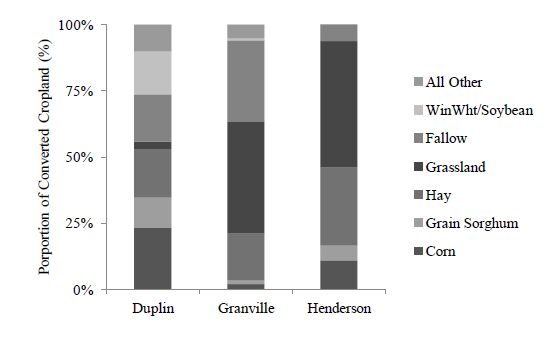
Figures(8)

Kevin R Caffrey, Mari S Chinn, Matthew W Veal. Biomass supply chain management in North Carolina (part 1): predictive model for cropland conversion to biomass feedstocks[J]. AIMS Energy, 2016, 4(2): 256-279. doi: 10.3934/energy.2016.2.256







 DownLoad:
DownLoad: