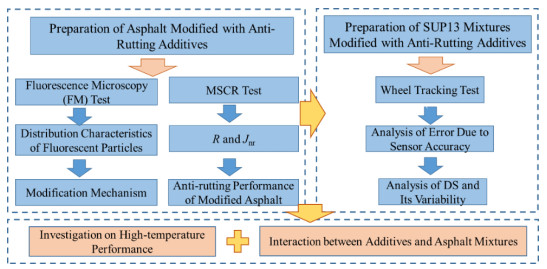
In order to investigate the asphalt mixture enhancement due to the granular anti-rutting additives, the multiple stress creep recovery (MSCR) test and wheel tracking test were performed to examine the effects of three anti-rutting additives on the high-temperature performance of binders and mixtures, respectively. Further, the interaction mechanism between anti-rutting additives and asphalt binder was revealed via a fluorescence microscopy (FM) test. The results indicate that the incorporation of anti-rutting additives causes a substantial increase in elasticity behavior for binders, along with a massive enhancement of dynamic stability for mixtures. Further, the enhancement of performance is not conclusively determined by the compatibility between the additive and asphalt. Instead, unmelted viscous-flow additives contribute to the enhancement of mixtures by interlocking, filling voids, cementing and wrapping the aggregate, which are essential mechanisms of mixture modification. This study contributes to selecting appropriate additives in engineering and enhancing anti-rutting additives based on their modification mechanism.
Citation: Gang Tian, Ying Gao, Conglin Chen, Qin Ye. Study on high-temperature performance evaluation of asphalt mixtures with anti-rutting additives[J]. Electronic Research Archive, 2024, 32(1): 160-173. doi: 10.3934/era.2024008
In order to investigate the asphalt mixture enhancement due to the granular anti-rutting additives, the multiple stress creep recovery (MSCR) test and wheel tracking test were performed to examine the effects of three anti-rutting additives on the high-temperature performance of binders and mixtures, respectively. Further, the interaction mechanism between anti-rutting additives and asphalt binder was revealed via a fluorescence microscopy (FM) test. The results indicate that the incorporation of anti-rutting additives causes a substantial increase in elasticity behavior for binders, along with a massive enhancement of dynamic stability for mixtures. Further, the enhancement of performance is not conclusively determined by the compatibility between the additive and asphalt. Instead, unmelted viscous-flow additives contribute to the enhancement of mixtures by interlocking, filling voids, cementing and wrapping the aggregate, which are essential mechanisms of mixture modification. This study contributes to selecting appropriate additives in engineering and enhancing anti-rutting additives based on their modification mechanism.

| [1] |
Y. Jia, S. Wang, J. Peng, Y. Gao, D. Hu, X. Zhao, Evaluation of pavement rutting based on driving safety of vehicles, Int. J. Pavement Res. Technol., 15 (2022), 457–469. https://doi.org/10.1007/s42947-021-00032-2 doi: 10.1007/s42947-021-00032-2

|
| [2] |
J. S. Tong, T. Ma, K. R. Shen, H. Y. Zhang, S. P. Wu, A criterion of asphalt pavement rutting based on the thermal-visco-elastic-plastic model, Int. J. Pavement Eng., 23 (2020), 4. https://doi.org/10.1080/10298436.2020.1792470 doi: 10.1080/10298436.2020.1792470
|
| [3] |
Y. Luan, Y. Ma, T. Ma, C. Wang, F. Xia, Research on the effects of asphalt performance on rutting resistance and its correlation with rutting performance indicators, Constr. Build. Mater., 400 (2023), 132773. https://doi.org/10.1016/j.conbuildmat.2023.132773 doi: 10.1016/j.conbuildmat.2023.132773
|
| [4] |
R. S. McDaniel, H. U. Bahia, Field evaluation of asphalt additives to control rutting and cracking, Transp. Res. Record, 1829 (2003), 47–54. https://doi.org/10.3141/1829-07 doi: 10.3141/1829-07
|
| [5] |
A. K. Ghuzlan, W. B. Al-Mistarehi, S. A. Al-Momani, Rutting performance of asphalt mixtures with gradations designed using Bailey and conventional Superpave methods, Constr. Build. Mater., 261 (2020), 119941. https://doi.org/10.1016/j.conbuildmat.2020.119941 doi: 10.1016/j.conbuildmat.2020.119941
|
| [6] |
J. Yang, J. Cui, J. Wan, X. Shi, Strategy of improving rutting resistance based on contribution rate of different layers, J. Southeast Univ. (Nat. Sci. Ed.), 37 (2007), 350–354. https://doi.org/10.3969/j.issn.1001-0505.2007.02.033 doi: 10.3969/j.issn.1001-0505.2007.02.033
|
| [7] | Y. L. Zhang, N. Li, Research on applicability of anti-rutting agent LY to different asphalt mixtures, in IOP Conference Series: Earth and Environmental Science, 587 (2020), 1. https://doi.org/10.1088/1755-1315/587/1/012029 |
| [8] | W. J. Wang, Design of Key Parameters and Study on the Performance of Bituminous Mixtures Mixed with Anti-Rutting Additive, M.S. thesis, Chang'an University, 2011. |
| [9] |
Z. Q. Zhang, Y. F. Luo, K. Zhang, Research on influence of different anti-rutting additives on high-temperature performance of asphalt mixture, J. Railw. Sci. Eng., 14 (2017), 964–970. https://doi.org/10.19713/j.cnki.43-1423/u.2017.05.012 doi: 10.19713/j.cnki.43-1423/u.2017.05.012
|
| [10] |
Y. S. Guan, H. Li, Z. X. Zhang, X. Y. Li, A new high rutting resistance asphalt mixture for intersection maintenance under heavy traffic, J. Test. Eval., 40 (2012), 1156–1161. https://doi.org/10.1520/JTE20120126 doi: 10.1520/JTE20120126
|
| [11] |
B. Wang, X. D. Hu, C. F. Cao, J. M. Wan, W. X. Gan, Z. W. Chen, et al., Development and characterization of permeative anti-rutting agent for asphalt mixture enhancement, Constr. Build. Mater., 364 (2023), 129937. https://doi.org/10.1016/j.conbuildmat.2022.129937 doi: 10.1016/j.conbuildmat.2022.129937
|
| [12] |
S. Han, W. Y. Ren, J. X. Jia, H. Yang, D. Y. Niu, Study on road performances of asphalt mixtures with granulated polymer anti-rutting additive, Int. J. Pavement Eng., 21 (2020), 257–265. https://doi.org/10.1080/10298436.2018.1461868 doi: 10.1080/10298436.2018.1461868
|
| [13] |
F. P. Xiao, D. F. Ma, J. Y. Wang, D. G. Cai, L. W. Lou, J. Yuan, Impacts of high modulus agent and anti-rutting agent on performances of airfield asphalt pavement, Constr. Build. Mater., 204 (2019), 1–9. https://doi.org/10.1016/j.conbuildmat.2019.01.138 doi: 10.1016/j.conbuildmat.2019.01.138
|
| [14] | JTG E20-2011, Standard Test Methods of Bitumen and Bituminous Mixtures for Highway Engineering, Ministry of Transport of the People's Republic of China, 2011. |
| [15] | JTG E42-2005, Test Methods of Aggregate for Highway Engineering, Ministry of Transport of the People's Republic of China, 2005. |
| [16] |
Z. Y. Wei, Y. S. Jia, S. Q. Wang, Z. R. Li, Y. S. Li, X. M. Wang, et al., Utilization of iron ore tailing as an alternative mineral filler in asphalt mastic: High-temperature performance and environmental aspects, J. Cleaner Prod., 335 (2022), 130318. https://doi.org/10.1016/j.jclepro.2021.130318 doi: 10.1016/j.jclepro.2021.130318
|
| [17] |
M. D. I. Domingos, A. L. Faxina, Susceptibility of asphalt binders to rutting: Literature review, J. Mater. Civ. Eng., 28 (2016), 04015134. https://doi.org/10.1061/(ASCE)MT.1943-5533.0001364 doi: 10.1061/(ASCE)MT.1943-5533.0001364
|










Figures(11) / Tables(4)
Gang Tian, Ying Gao, Conglin Chen, Qin Ye. Study on high-temperature performance evaluation of asphalt mixtures with anti-rutting additives[J]. Electronic Research Archive, 2024, 32(1): 160-173. doi: 10.3934/era.2024008







 DownLoad:
DownLoad: